Стоит ли вступать в жилищный накопительный кооператив: мнение адвоката
Организация жилищного накопительного кооператива – один из способов привлечь деньги граждан на строительство или покупку жилья. Однако прежде, чем вступать в дружные ряды членов ЖНК, стоит поинтересоваться, а так ли прекрасен этот институт продажи жилья как его «малюют».
Тактика формирования
Вступление граждан в жилищный накопительный кооператив и функционирование ЖНК регулируются Жилищным кодексом РФ и законом №215 «О жилищных накопительных кооперативах» от 30 декабря 2004 года.
ЖНК, как и любая организация, должен быть зарегистрирован налоговой службой в Едином государственном реестре юридических лиц (далее ЕГРЮЛ). Только с момента регистрации кооператив считается созданным. Документы на регистрацию кооператива подает председатель правления или иное уполномоченное собранием кооператива лицо.
Список документов не велик: заявление о регистрации, решение о создании ЖНК, устав и квитанция об оплате пошлины.
Для создания кооператива предусмотрено минимальное количество его участников – 50 человек (максимальное число членов – 50 тысяч) и соответствующее их решение о создании ЖНК.
Структура и управление
ЖНК имеет свою собственную структуру правления, чьими органами являются общее собрание, правление, ревизионная комиссия и исполнительные органы (например, директор).
Общее собрание является высшим органом ЖНК и правомочно решать абсолютно все вопросы кооператива.
Решения на общих собраниях считаются принятыми, если за них проголосовало более половины участвующих в собрании. В исключительных случаях, в частности, при утверждении устава кооператива или принятии решения о ликвидации кооператива, легитимность решений будет основана уже при голосовании «за» двух третей участников общего собрания, составляющих более половины членов кооператива.
В порядке необщей очереди
Существование ЖНК поддерживается вступительными, членскими и дополнительными взносами. Аккумулирование средств членов ЖНК для приобретения или строительства жилья осуществляется за счет паевых взносов и заемных (кредитных) средств. Кредитные средства поступают на расчетный счет кооператива, и отдельные его члены не вправе распоряжаться полученным займом: деньги расходуются на основании решения общего собрания и в целях, установленных им.
Аккумулирование средств членов ЖНК для приобретения или строительства жилья осуществляется за счет паевых взносов и заемных (кредитных) средств. Кредитные средства поступают на расчетный счет кооператива, и отдельные его члены не вправе распоряжаться полученным займом: деньги расходуются на основании решения общего собрания и в целях, установленных им.
Право на получение квартиры у члена ЖНК возникает с момента внесения части паевого взноса, определенной в уставе кооператива, в порядке очередности, установленной уставом или решением общего собрания, и с учетом размеров и сроков внесенного пая.
Передача квартиры члену ЖНК осуществляется после оформления ее в собственность кооператива по решению органа правления кооператива.
До полной выплаты пая член ЖНК может только пользоваться помещением, а после полной выплаты помещение оформляется в его собственность на основании справки о выплате пая.
В чем подвох
Существует несколько поводов хорошенько подумать, прежде чем вступать в члены ЖНК.
Во-первых, у договоров, которые оформляются через ЖНК, отсутствует государственная регистрация, что нередко приводит к случаям двойных продаж, когда владельцами одной и той же жилплощади могут стать несколько человек.
Во-вторых, жилищный накопительный кооператив – это единственная форма организации, которая по закону (статья 110 Жилищного кодекса РФ) вправе требовать от своих членов дополнительные средства на достройку дома по уже оплаченным договорам. При этом такие дополнительные взносы могут в разы превышать сумму первоначального взноса.
Кроме того, при вступлении в ЖНК новые члены обязаны оплатить вступительные, членские, целевые, дополнительные и иные взносы, которые в большинстве своем не возвращаются в случае выхода человека из кооператива. Возврату подлежат лишь целевые взносы на строительство или покупку недвижимости. Впрочем, и в этом случае бывший член кооператива столкнется с целым рядом трудностей. Во-первых, в уставе ЖНК могут быть прописаны сроки, по истечении которых и производится возврат целевых взносов.
И, наконец, еще один подвох: поскольку ЖНК является некоммерческой структурой (а по факту — простым собранием граждан), то и сфера его ответственности минимальна. Финансовые обязательства ЖНК ограничиваются суммой внесенных самими пайщиками средств. Кроме того, в договорах кооперативов редко прописываются четкие сроки предоставления жилья пайщикам, поэтому последние не застрахованы и не защищены законом от просрочек и связанных с этим убытков. Получается, у члена ЖНК нет никаких гарантий того, что он получит свое жилье вовремя, не переплатив за него вдвое. В случае же возможных убытков и неустоек предъявлять претензии будет некому.
Олег Сухов, адвокат, президент «Гильдии юристов рынка недвижимости»
Статья 3. Основные положения о жилищных накопительных кооперативах / КонсультантПлюс
Статья 3. Основные положения о жилищных накопительных кооперативах
1. За исключением деятельности, предусмотренной настоящим Федеральным законом, иную деятельность жилищный накопительный кооператив осуществлять не вправе.
За исключением деятельности, предусмотренной настоящим Федеральным законом, иную деятельность жилищный накопительный кооператив осуществлять не вправе.
2. Наименование жилищного накопительного кооператива должно содержать слова «жилищный накопительный кооператив». Юридические лица, не отвечающие требованиям настоящего Федерального закона, не вправе использовать в своих наименованиях слова «жилищный накопительный кооператив».
3. Кооператив имеет в собственности обособленное имущество, учитываемое на его самостоятельном балансе, от своего имени может приобретать и осуществлять имущественные и личные неимущественные права, нести обязанности, быть истцом и ответчиком в суде. Имущество кооператива образуется за счет паевых и иных взносов членов кооператива, доходов, полученных кооперативом от осуществляемой им предпринимательской деятельности, которая служит достижению целей, ради которых кооператив создан, и соответствует этим целям, добровольных пожертвований и иных не запрещенных законом источников.
4. Кооператив считается созданным как юридическое лицо со дня внесения соответствующей записи в единый государственный реестр юридических лиц. Кооператив создается без ограничения срока деятельности, если иное не установлено его уставом.
5. Отношения между кооперативом и его членами возникают на основании членства в кооперативе в соответствии с требованиями Гражданского кодекса Российской Федерации, настоящего Федерального закона, принятых в соответствии с ним нормативных правовых актов Российской Федерации, нормативных актов Центрального банка Российской Федерации (далее — Банк России), устава кооператива, а также в соответствии с решениями органов кооператива, принятыми в пределах их компетенции.
(в ред. Федерального закона от 23.07.2013 N 251-ФЗ)
6. Кооператив не может состоять в договорных отношениях с членами кооператива, в результате которых устанавливаются, изменяются или прекращаются гражданские права и обязанности, связанные с осуществлением деятельности кооператива по привлечению и использованию денежных средств граждан на приобретение жилых помещений.
7. Кооператив в установленном порядке вправе открывать счета в банках, находящихся на территории Российской Федерации.
8. Кооператив должен иметь круглую печать, содержащую его полное наименование на русском языке и указание на место нахождения кооператива. Печать кооператива может содержать также его фирменное наименование на любом языке народов Российской Федерации и (или) иностранном языке. Кооператив вправе иметь штампы и бланки со своим наименованием, собственную эмблему и другие средства индивидуализации.
9. Кооператив обязан обеспечить возможность предоставления в Банк России электронных документов, а также возможность получения от Банка России электронных документов в порядке, установленном Банком России.
(часть 9 введена Федеральным законом от 13.07.2015 N 231-ФЗ)
Открыть полный текст документа
Жилищно-строительные и жилищно-накопительные кооперативы | Народный вопрос.РФ
Жилищно-строительные и жилищно-накопительные кооперативы
Помимо покупки жилья на вторичном рынке и покупки квартиры в уже выстроенном доме, процедуру которых регулирует ГК РФ, граждане могут приобрести недвижимость и иными способами, одним из которых является участие в жилищном кооперативе (ЖК) или жилищно-строительном кооперативе (ЖСК) — разновидностях потребительских кооперативов.
Члены жилищного кооператива своими средствами участвуют в приобретении, реконструкции и последующем содержании многоквартирного дома. При проведении реконструкции многоквартирного дома жилищный кооператив в соответствии с законодательством о градостроительной деятельности выступает в качестве застройщика и обеспечивает на принадлежащем ему земельном участке реконструкцию этого дома в соответствии с выданным такому кооперативу разрешением на строительство.
Члены жилищно-строительного кооператива своими средствами участвуют в строительстве, реконструкции и последующем содержании многоквартирного дома. Жилищно-строительный кооператив в соответствии с законодательством о градостроительной деятельности выступает в качестве застройщика и обеспечивает на принадлежащем ему земельном участке строительство, реконструкцию многоквартирного дома в соответствии с выданным такому кооперативу разрешением на строительство. Жилищно-строительный кооператив, за исключением жилищно-строительных кооперативов, создание которых предусмотрено Федеральным законом от 24 июля 2008 года N 161-ФЗ «О содействии развитию жилищного строительства», не вправе осуществлять одновременно строительство более одного многоквартирного дома с количеством этажей более чем 3.
Жилищный накопительный кооператив — потребительский кооператив, созданный как добровольное объединение граждан на основе членства в целях удовлетворения потребностей членов кооператива в жилых помещениях путем объединения членами кооператива паевых взносов.
Членство в жилищном кооперативе возможно с 16 лет. Человек, вступивший в него приобретает право на имущественный пай — т. е. одну энную от общей площади предполагаемого дома. Количество членов жилищного кооператива не может быть менее чем пять, но не должно превышать количество жилых помещений в строящемся или приобретаемом кооперативом многоквартирном доме. Порядок и условия внесения паевого взноса членом жилищного кооператива определяются уставом жилищного кооператива. Заявление о приеме в члены жилищного кооператива должно быть рассмотрено в течение месяца правлением жилищного кооператива и утверждено решением общего собрания членов жилищного кооператива (конференции). Гражданин или юридическое лицо признается членом жилищного кооператива с момента уплаты вступительного взноса после утверждения решения о приеме в члены жилищного кооператива общим собранием членов кооператива (конференцией).
Члену жилищного кооператива, не выплатившему полностью паевого взноса и исключенному из жилищного кооператива, выплачивается сумма его пая в сроки и на условиях, которые предусмотрены уставом кооператива. Основанием владения, пользования и в установленных законодательством пределах распоряжения жилым помещением является членство в жилищном кооперативе. После выплаты взноса он приобретает права и на общую площадь, имеющуюся в кооперативе.
Жилищный кооператив по решению общего собрания его членов (конференции) может быть преобразован в товарищество собственников жилья.
ЖСК как организационно-правовую форму объединения граждан для строительства используют обманутые дольщики, желающие достроить своё жильё после того, как строительные компании, привлекавшие их средства, прекращают строительство. ЖСК могут сами выступать как застройщики, а могут участвовать в качестве инвестора или участника долевого строительства.
ЖСК могут сами выступать как застройщики, а могут участвовать в качестве инвестора или участника долевого строительства.
Жилищно-накопительный кооператив действует на основании положений Федерального закона от 30.12.2004 N 215-ФЗ «О жилищных накопительных кооперативах», а не глав 11-12 Жилищного кодекса. Число членов кооператива не может быть менее чем 50 человек и более чем 5 000 человек. Членство в кооперативе возникает после внесения соответствующих сведений о гражданине, подавшем заявление о приеме в члены кооператива, в единый государственный реестр юридических лиц в порядке, предусмотренном настоящим Федеральным законом и Федеральным законом от 8 августа 2001 года N 129-ФЗ «О государственной регистрации юридических лиц и индивидуальных предпринимателей», со дня внесения гражданином вступительного членского взноса и первого платежа в счет паевого взноса — в ЖСК.
В случае невнесения гражданином вступительного членского взноса или первого платежа в счет паевого взноса в течение трех месяцев со дня внесения соответствующих сведений в единый государственный реестр юридических лиц единоличный исполнительный орган кооператива обязан направить в федеральный орган исполнительной власти, уполномоченный на осуществление государственной регистрации юридических лиц, заявление об аннулировании сведений о гражданине, подавшем заявление о приеме в члены кооператива, в едином государственном реестре юридических лиц. Исключение из членов кооператива возможно только в судебном порядке, а не лишь по решению общего собрания.
Исключение из членов кооператива возможно только в судебном порядке, а не лишь по решению общего собрания.
То есть порядок регистрации членства в ЖНК более строгий и четкий, чем в ЖК или ЖСК.
Деятельность ЖНК вообще более четко и подробно прописана в законе, чем деятельность ЖК или ЖСК, поэтому ЖНК выглядит более безопасной формой вложения средств в приобретение недвижимости. В законе, в частности, указано, что член кооператива или другие лица, имеющие право на пай, внесшие в полном размере паевой взнос за жилое помещение, переданное кооперативом в пользование члену кооператива, приобретают право собственности на это жилое помещение. Кооператив обязан передать члену кооператива или другим лицам, имеющим право на пай, указанное жилое помещение свободным от каких-либо обязательств, в то время как член ЖСК по прежнему будет владеть только лишь паем.
Кроме того ЖНК должны соблюдать финансовые нормативы, свидетельствующие о их финансовой устойчивости.
И ЖСК и ЖНК предусматривают наследование пая, возможно сдачи жилья в наем, временного проживания в нем третьих лиц, перехода пая от одного члена к другому.
Следует помнить что ЖСК и ЖНК — это не форма покупки квартиры, а форма инвестирования в недвижимость с соответствующими рисками.
ЖНК что это такое, схема работы жнк
Жилищный накопительный кооператив «Доступные Метры» создан в соответствии с Федеральным Законом № 215-ФЗ «О жилищных накопительных кооперативах» от 30.12.2004г.
Главная цель ЖНК «Доступные Метры» — помощь в приобретении жилых помещений всем, кому требуется улучшение жилищных условий, причем быстрое, надежное и недорогое.
Основные преимущества приобретения квартиры (дома) в ЖНК «Доступные метры»:
ЖНК подбирает, осуществляет юридическую проверку всех документов и приобретает для члена кооператива только ту квартиру или дом, который выбирает сам член кооператива.

Возможность уменьшить расходы на приобретение квартиры по сравнению с ипотечным и другими видами кредитования.
Возможность приобрести квартиру с ремонтом, сделанным в соответствии с пожеланиями члена кооператива. При этом оплата ремонта осуществляется с рассрочкой платежа.
Каждый человек сам выбирает оптимальный вариант оплаты стоимости квартиры(дома) исходя из своих возможностей и потребностей.
Возможность значительно сократить срок приобретения квартиры по сравнению с ипотечными продуктами банков.
Все доходы ЖНК распределяются между пайщиками пропорционально паям. Таким образом накопление Пая осуществляется быстрее.

Ваши платежи напрямую зависят от Вашего выбора и Ваших возможностей, так как рассчитываются исходя из стоимости выбранного Вами и планируемого к приобретению жилого помещения в соответствии с выбранной Вами формой участия в деятельности ЖНК «Доступные Метры».
В первую очередь, Вы должны определить три важных ориентира:
какую квартиру (по стоимости и другим характеристикам) Вы планируете приобрести
какую сумму и за какой период необходимо накопить для приобретения выбранной квартиры
определить посильный размер ежемесячных платежей для приобретения жилья
Методика приобретения жилья в ЖНК «Доступные Метры»
Формы участия в ЖНК Доступные метры:
Вы прагматичный человек и готовы накопить 50% от стоимости желаемого жилого помещения, остальные денежные средства на приобретение Вам выделит ЖНК в рассрочку под 3,5% годовых. Причем погашать вы будете дифференцированными платежами. Ведь рассрочка выгодней ипотеки!
Причем погашать вы будете дифференцированными платежами. Ведь рассрочка выгодней ипотеки!
Не откладывайте! Начинайте копить уже сейчас. Ваша переплата будет в разы меньше, чем по ипотечному кредиту. Убедитесь сами — закажите расчет!
Вы расчетливый человек и ставки по ипотечным кредитам Вас не устраивают. Воспользовавшись данной формой участия в ЖНК «Доступные Метры», вы уменьшите переплату по сравнению с ипотечным кредитом.
Смотрите сами: Вы накопили 30% от стоимости жилого помещения ЖНК Вам выделяет еще 30% под 3,5% годовых и банк-партнер предоставляет оставшиеся средства для приобретения в соответствии с условиями ипотечного кредита.
Вам понравилась квартира на вторичном рынке, вы хотите сменить район, расширить домашнее пространство или разъехаться и в счет оплаты вы готовы сдать вашу квартиру.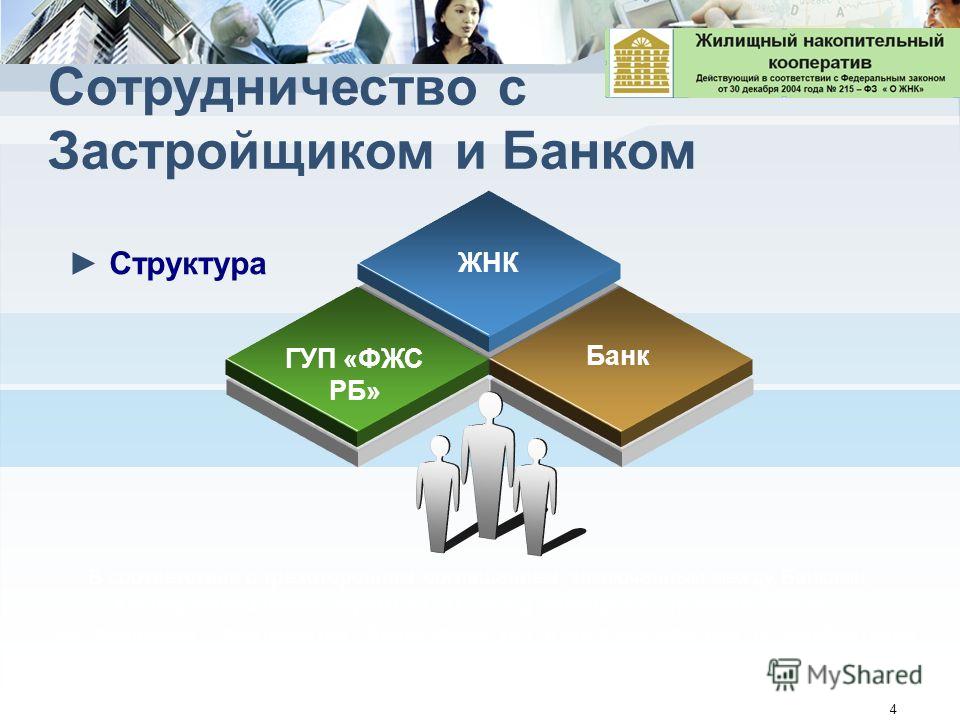 Вас останавливает только недостаточность средств для осуществления задуманных планов.
Вас останавливает только недостаточность средств для осуществления задуманных планов.
Вы можете воспользоваться данной формой участия. Доплату вам выделит ЖНК в рассрочку под 3,5% годовых. Не оттягивайте осуществление ваших планов. Звоните!
Записаться на консультациюВы хотите приобрести новостройку. Кооператив предоставит вам эту возможность, примет в счет оплаты вашу квартиру. При этом вы будете проживать в ней, до заселения в новую. А недостающие денежные средства для оплаты нового предоставит в рассрочку под 3,5% годовых.
Обновляетесь, это будет новый этап в вашей жизни!
Записаться на консультациюВам понравилась квартира из вариантов, предлагаемых ЖНК. И вы в счет оплаты готовы внести свою. Тогда данная форма участия для вас!
И вы в счет оплаты готовы внести свою. Тогда данная форма участия для вас!
Не ждите «у моря погоды»! Желающие на оптимальный вариант всегда найдутся!
Записаться на консультацию| Имя | Доля в уставном капитале | |||
|---|---|---|---|---|
| Галимарданова Данфира Газизяновна | Физическое лицо | ИНН 165007782371 | ||
| Дорина Наталья Николаевна | Физическое лицо | ИНН 667116792514 | ||
| Кузьмина Галина Федоровна | Физическое лицо | ИНН 263507692125 | ||
| Мочалина Наталья Витальевна | Физическое лицо | ИНН 522300732245 | ||
| Ракова Галина Викторовна | Физическое лицо | ИНН 732800457310 | ||
| Елизова Валентина Анатольевна | Физическое лицо | ИНН 281001513469 | ||
| Потанин Артем Николаевич | Физическое лицо | ИНН 027620724136 | ||
| Казарян Лусине Грачиковна | Физическое лицо | ИНН 333400812987 | ||
| Сакерина Галина Владимировна | Физическое лицо | ИНН 243500338845 | ||
| Беляк Елена Ивановна | Физическое лицо | ИНН 343500267883 | ||
| Курченко Валерия Николаевна | Физическое лицо | ИНН 424300826536 | ||
| Короткова Елена Юрьевна | Физическое лицо | ИНН 501710170919 | ||
| Курбанова Раиля Рифмировна | Физическое лицо | ИНН 860904251985 | ||
| Михайлова Евгения Леонидовна | Физическое лицо | ИНН 471206757659 | ||
| Кадочникова Евгения Викторовна | Физическое лицо | ИНН 590772208004 | ||
| Парфенов Олег Николаевич | Физическое лицо | ИНН 352801959900 | ||
| Углова Ирина Павловна | Физическое лицо | ИНН 246506254650 | ||
| Парфенова Ольга Николаевна | Физическое лицо | ИНН 352811734528 | ||
| Лоскутов Евгений Дмитриевич | Физическое лицо | ИНН 164811129395 | ||
| Яковлева Олеся Петровна | Физическое лицо | ИНН 420590727270 | ||
| Султанов Эдуард Анварович | Физическое лицо | ИНН 021301623412 | ||
| Касьянова Екатерина Федоровна | Физическое лицо | ИНН 590421540965 | ||
| Каримуллина Эльвира Эдуардовна | Физическое лицо | ИНН 165905824050 | ||
| Султанова Любовь Александровна | Физическое лицо | ИНН 025000749031 | ||
| Еруков Михаил Геннадьевич | Физическое лицо | ИНН 246213725941 | ||
| Кузякина Марина Евгеньевна | Физическое лицо | ИНН 244603093109 | ||
| Волкова Наталья Владимировна | Физическое лицо | ИНН 245206350859 | ||
| Благодатная Мария Олеговна | Физическое лицо | ИНН 667109555583 | ||
| Маслянка Элеоноры Викторовны | Физическое лицо | ИНН 667009148986 | ||
| Туктагулова Алия Галимзяновна | Физическое лицо | ИНН 560502482649 | ||
| Кузюков Сергей Сергеевич | Физическое лицо | ИНН 110305456100 | ||
| Валиева Лилия Дамировна | Физическое лицо | ИНН 165028483608 | ||
| Шаяхметова Карина Рафаилевна | Физическое лицо | ИНН 161404957849 | ||
| Казаков Роман Иванович | Физическое лицо | ИНН 163204577498 | ||
| Галиева Лилия Филаритовна | Физическое лицо | ИНН 026906534622 | ||
| Низамиев Рамиль Рустамович | Физическое лицо | ИНН 160903676961 | ||
| Копотилова Екатерина Сергеевна | Физическое лицо | ИНН 161404878643 | ||
| Гафарова Анна Андреевна | Физическое лицо | ИНН 161604998049 | ||
| Курманов Роман Геннадьевич | Физическое лицо | ИНН 745111227480 | ||
| Барыкина Амалия Владимировна | Физическое лицо | ИНН 301612185291 | ||
| Степанова Оксана Николаевна | Физическое лицо | ИНН 470321027722 | ||
| Демина Анастасия Александровна | Физическое лицо | ИНН 760409623770 | ||
| Смородина Юлия Валерьевна | Физическое лицо | ИНН 616510464927 | ||
| Тишкова Юлия Сергеевна | Физическое лицо | ИНН 623000129545 | ||
| Хлебушкин Лев Вячеславович | Физическое лицо | ИНН 780438659855 | ||
| Ильина Наталия Викторовна | Физическое лицо | ИНН 110115309852 | ||
| Хафизова Ирина Ивановна | Физическое лицо | ИНН 165903827271 | ||
| Алимова Ландыш Завдятовна | Физическое лицо | ИНН 165915875400 | ||
| Салимуллин Малик Альбертович | Физическое лицо | ИНН 165129441900 | ||
| Попов Максим Сергеевич | Физическое лицо | ИНН 751503536658 | ||
| Гончар Альбина Витальевна | Физическое лицо | ИНН 740800019478 | ||
| Гребнева Инна Викторовна | Физическое лицо | ИНН 742400975730 | ||
| Кошкарева Татьяна Николаевна | Физическое лицо | ИНН 424303750966 | ||
| Сафина Эльвина Насиповна | Физическое лицо | ИНН 166026842998 |
В Курганской области решается вопрос об обеспечении граждан доступным жильем | Новости Кургана и Курганской области
Может ли жилищно-накопительный кооператив стать решением проблемы?
В Курганской области актуальным остается вопрос об обеспечении граждан доступным жильем. По оценкам Банка России лишь 35% семей могут позволить себе ипотеку при покупке квартиры из расчета 18 квадратных метров жилой площади на человека.
По оценкам Банка России лишь 35% семей могут позволить себе ипотеку при покупке квартиры из расчета 18 квадратных метров жилой площади на человека.
В Указе Президента Российской Федерации от 7 мая 2018 года, определяющем цели развития России до 2024 года, говорится о том, что к 2024 году необходимо обеспечить доступным жильем семьи со средним достатком. Ежегодно должны улучшаться жилищные условия, по меньшей мере, пяти миллионов российских семей. Согласно национальному проекту «Жилье и городская среда» объемы жилищного строительства к 2024 году должны увеличиться до 120 миллионов квадратных метров в год. Это означает, что необходимо искать дополнительные способы финансирования жилищного строительства.
На совместном заседании Совета по улучшению инвестиционного климата в городе Кургане и Совета по вопросам развития малого и среднего предпринимательства и конкуренции при администрации города Кургана рассматривался вопрос о целесообразности внедрения в регионе института жилищной накопительной кооперации.
Сегодня жилищные накопительные кооперативы (ЖНК) – это альтернативный способ накопления денежных средств для приобретения жилой недвижимости в собственность.
«Целевую модель устойчивого действующего жилищного накопительного кооператива разработал Банк России, – пояснил Евгений Шлепенков, заместитель управляющего отделением по Курганской области Уральского главного управления Центрального банка Российской Федерации. – ЖНК помогут обеспечить жильем граждан, наиболее остро нуждающихся в улучшении жилищных условий. ЖНК привлекают денежные средства пайщиков для приобретения недвижимости и затем передают ее пайщику после выплаты им полной стоимости пая кооперативу. Специализированному оператору-застройщику предоставляется земельный участок для застройки на льготных условиях и выделяется необходимая для строительства инфраструктура. Строительство осуществляется в течение 1-2 лет, граждане получают квартиры в пользование в течение 1,5-2,5 лет в зависимости от срока накопления первоначального взноса. Первоначальный взнос – это часть паевого взноса, после ее внесения у кооператива возникает право на приобретение (строительство) для члена ЖНК жилого помещения в соответствии с положениями Федерального закона «О жилищных накопительных кооперативах» № 215-ФЗ».
Первоначальный взнос – это часть паевого взноса, после ее внесения у кооператива возникает право на приобретение (строительство) для члена ЖНК жилого помещения в соответствии с положениями Федерального закона «О жилищных накопительных кооперативах» № 215-ФЗ».
Получив квартиру в пользование, пайщик может проживать в квартире, регистрироваться в ней, сдавать ее в аренду. Однако жилье остается в собственности кооператива. Далее член кооператива продолжает вносить членские взносы для погашения оставшейся стоимости жилья. После полной выплаты стоимости жилья оно переходит из собственности кооператива в собственность члена кооператива.
Преимущества модели ЖНК – переплата по приобретению жилья составляет 2-3% годовых, контроль стоимости квадратного метра отстраиваемого жилья (эконом-класса) позволяет удерживать ее значение в пределах цен, устанавливаемых Министерством строительства Российской Федерации. Строительство контролируется государством, устойчивость ЖНК контролируется Банком России.
Было предложено создать рабочую группу, чтобы проанализировать целесообразность внедрения института ЖНК в Кургане и Курганской области. Предполагается, что в рабочую группу войдут представители Курганской ипотечно-жилищной корпорации, администрации города Кургана, отделения по Курганской области Уральского главного управления Центрального банка Российской Федерации, депутаты городской Думы, представители строительных организаций, действующих на территории региона.
Если вы стали свидетелем интересного события, присылайте сообщения, фото и видео в Viber и WhatsApp по номеру тел. : +79195740453, в нашей группе «В Контакте»
Базовые истины | Фирма «Культбытстрой»
Основой деятельности ЖНК является принцип взаимной финансовой поддержки или, другими словами, принцип кассы взаимопомощи.
ЖНК — это некоммерческая организация. Каждый член кооператива обеспечивается жильем за счет собственных средств и средств участников ЖНК, поступающих в общий паевой фонд. Вселиться в квартиру Вы сможете, накопив часть стоимости жилья. Недостающие средства будут добавлены из паевого фонда ЖНК (но не более накопленной суммы). Предоставленные средства Вы должны вернуть в паевой фонд в соответствии с графиком рассрочки платежей в срок, установленный формой участия. В результате каждый член ЖНК выплачивает полную стоимость квартиры, но в рассрочку.
Вселиться в квартиру Вы сможете, накопив часть стоимости жилья. Недостающие средства будут добавлены из паевого фонда ЖНК (но не более накопленной суммы). Предоставленные средства Вы должны вернуть в паевой фонд в соответствии с графиком рассрочки платежей в срок, установленный формой участия. В результате каждый член ЖНК выплачивает полную стоимость квартиры, но в рассрочку.
Все средства общего паевого фонда вкладываются исключительно в недвижимость, которая приобретается для членов кооператива. Квартира приобретается кооперативом, когда пайщиком уплачена сумма, предусмотренная избранной им формой участия, но не менее 50-ти процентов от стоимости квартиры на момент её приобретения.
Член ЖНК и члены его семьи проживают в приобретенной для них квартире, оплачивают коммунальные услуги, оформляют на себя телефон и т.п. В общем, живут полноценной жизнью, выплачивая членские и паевые взносы. После выплаты пая в полном объёме и ввода дома в эксплуатацию (при долевом участии в строительстве жилья), член ЖНК, в соответствии с ГК РФ (ст. 218, п.4) становится полноправным собственником приобретенной для него квартиры. Свидетельство о праве собственности на квартиру пайщик получает после регистрации права собственности кооперативом, обратившись в регистрирующий орган вместе с представителем кооператива, лишь предоставляя справку о выплате пая.
218, п.4) становится полноправным собственником приобретенной для него квартиры. Свидетельство о праве собственности на квартиру пайщик получает после регистрации права собственности кооперативом, обратившись в регистрирующий орган вместе с представителем кооператива, лишь предоставляя справку о выплате пая.
Стоимость недвижимости фиксируется на момент ее приобретения для члена ЖНК и размер выплат не увеличивается в связи с ростом ее цены. Это – еще одно преимущество представленной системы.
Принципиальное отличие ЖНК от жилищных и жилищно-строительных кооперативов заключается в регистрации всех членов кооператива в специальном реестре, который ведет налоговый орган. Кроме этого, жилье может быть подобрано не в каком-то конкретном доме (месте), а в любом интересующем члена кооператива доме или вложено в строительство строящегося дома (квартиры). И наконец, в качестве паевого взноса могут выступать предоставленные члену кооператива субсидии и субвенции.
Средства из Паевого фонда Кооператива могут расходоваться только на приобретение недвижимости для членов ЖНК. Они аккумулируются на отдельном расчетном счете. Текущая деятельность ЖНК — содержание офиса, рекламные затраты, оплата труда сотрудников и развитие в целом, осуществляется за счет вступительных и членских взносов.
Они аккумулируются на отдельном расчетном счете. Текущая деятельность ЖНК — содержание офиса, рекламные затраты, оплата труда сотрудников и развитие в целом, осуществляется за счет вступительных и членских взносов.
Достижения в области передачи сигналов WNK о метаболизме соли и калия: клинические последствия — Полный текст — Американский журнал нефрологии 2012, Vol. 35, № 4
Аннотация
Недавние данные, полученные в связи с открытием семейства киназ, вовлеченных в артериальную гипертензию, теперь указывают на лежащие в основе молекулярные механизмы, определяющие Na + , K + и обращение с водой в нефроне. Эти новые ключевые игроки необходимо понимать, чтобы полностью понять патофизиологию, проявления и лечение распространенных клинических состояний, таких как гиповолемический шок, застойная сердечная недостаточность, первичный гиперальдостеронизм, нефротический синдром и артериальная гипертензия. Именно посредством анализа состояния объема и электролитных нарушений, которые обычно проявляются при этих заболеваниях, мы можем начать создавать связь между абстрактной концепцией регуляции киназы и тем, как пациент будет реагировать на конкретное лечение. Этот обзор является попыткой восполнить этот пробел.
Именно посредством анализа состояния объема и электролитных нарушений, которые обычно проявляются при этих заболеваниях, мы можем начать создавать связь между абстрактной концепцией регуляции киназы и тем, как пациент будет реагировать на конкретное лечение. Этот обзор является попыткой восполнить этот пробел.
© 2012 S. Karger AG, Базель
Введение
Баланс Na + и K + включает внутриклеточное и внеклеточное распределение этих ионов.Na + /K + АТФаза распределяет эти ионы таким образом, что Na + является наиболее распространенным внеклеточным ионом, а K + является наиболее распространенным внутриклеточным ионом. Колебания Na + и воды влияют на состояние объема. Низкое потребление натрия активирует механизмы удержания объема, необходимые для поддержания внеклеточного объема, в то время как избыток соли и воды в физиологических условиях выводится за счет натрийуреза под давлением, а в патологических ситуациях приводит к образованию отеков. Напротив, распределение K + находится в прямой зависимости от потребления с пищей, внутриклеточной обработки и скорости потока мочи. Прием пищи, как правило, не является причиной изменения K + в плазме. Иногда изменение обращения клеток с K + (например, рабдомиолиз) может привести к гиперкалиемии, развитию сердечных аритмий и смерти. Скорость потока мочи, однако, является основным фактором, определяющим способность почек успешно экскретировать K + таким образом, что высокая скорость потока мочи способствует экскреции K + , а низкая скорость потока мочи приводит к задержке K + .То, как недавно открытые киназы без лизина (K) (WNK) участвуют в регуляции систем транспорта ионов в дистальном отделе нефрона, стало областью интенсивных исследований в последние годы [обширные обзоры см. [1,2] ].
Напротив, распределение K + находится в прямой зависимости от потребления с пищей, внутриклеточной обработки и скорости потока мочи. Прием пищи, как правило, не является причиной изменения K + в плазме. Иногда изменение обращения клеток с K + (например, рабдомиолиз) может привести к гиперкалиемии, развитию сердечных аритмий и смерти. Скорость потока мочи, однако, является основным фактором, определяющим способность почек успешно экскретировать K + таким образом, что высокая скорость потока мочи способствует экскреции K + , а низкая скорость потока мочи приводит к задержке K + .То, как недавно открытые киназы без лизина (K) (WNK) участвуют в регуляции систем транспорта ионов в дистальном отделе нефрона, стало областью интенсивных исследований в последние годы [обширные обзоры см. [1,2] ].
WNK представляют собой группу серин/треонинкиназ, которые связаны с развитием гипертонии у людей [3]. Интронные делеции гена WNK1, приводящие к гиперэкспрессии нормального в остальном WNK1, или миссенс-мутации гена WNK4 в высококонсервативной кислой области являются причиной наследственной формы солечувствительной гипертензии, известной как псевдогипоальдостеронизм II типа (PHA II; также известный как семейная гиперкалиемическая гипертензия или синдром Гордона) [3]. Это редкое аутосомно-доминантное заболевание, характеризующееся артериальной гипертензией, гиперкалиемией и метаболическим ацидозом [4,5]. Клиническая картина PHA II является точным зеркальным отражением болезни Гительмана, аутосомно-рецессивного синдрома, характеризующегося гипокалиемическим метаболическим алкалозом с артериальной гипотензией, что обусловлено инактивирующими мутациями почечного тиазид-чувствительного Na-Cl-котранспортера (NCC) [6,7]. ] расположены в дистальном извитом канальце (DCT) [8,9,10]. Тот факт, что эти синдромы отражают друг друга, и клиническое наблюдение, что PHA II может быть скорректировано низкими дозами тиазидов [4], привели к предположению, что модуляция NCC с помощью WNK может лежать в основе патофизиологии PHA II [11]. .Наблюдения за последние несколько лет подтвердили эту гипотезу и выявили важную роль WNK, которая не только помогает объяснить PHA II, но также помогает прояснить молекулярную природу обработки соли и / или калия в физиологических условиях и при общих клинических синдромах, таких как застойное сердце.
Это редкое аутосомно-доминантное заболевание, характеризующееся артериальной гипертензией, гиперкалиемией и метаболическим ацидозом [4,5]. Клиническая картина PHA II является точным зеркальным отражением болезни Гительмана, аутосомно-рецессивного синдрома, характеризующегося гипокалиемическим метаболическим алкалозом с артериальной гипотензией, что обусловлено инактивирующими мутациями почечного тиазид-чувствительного Na-Cl-котранспортера (NCC) [6,7]. ] расположены в дистальном извитом канальце (DCT) [8,9,10]. Тот факт, что эти синдромы отражают друг друга, и клиническое наблюдение, что PHA II может быть скорректировано низкими дозами тиазидов [4], привели к предположению, что модуляция NCC с помощью WNK может лежать в основе патофизиологии PHA II [11]. .Наблюдения за последние несколько лет подтвердили эту гипотезу и выявили важную роль WNK, которая не только помогает объяснить PHA II, но также помогает прояснить молекулярную природу обработки соли и / или калия в физиологических условиях и при общих клинических синдромах, таких как застойное сердце. недостаточность, гиповолемия, первичный гиперальдостеронизм и гиперкалиемия.
недостаточность, гиповолемия, первичный гиперальдостеронизм и гиперкалиемия.
Минералокортикоидный гормон альдостерон в течение многих лет считается ключевым игроком в модуляции транспортных систем Na + и K + в дистальных отделах нефрона.Благодаря открытию киназ WNK только недавно начали выяснять точный механизм, с помощью которого почки способны осуществлять различные реакции на гиперкалиемию или гиповолемию, состояния, при которых альдостерон повышен. Альдостерон секретируется, когда необходимо удерживать соль (диета с низким содержанием соли/гиповолемия) или когда необходимо выводить калий (диета с высоким содержанием калия/гиперкалиемия). Во время истощения объема реабсорбция соли увеличивается, но секреция калия остается неизменной, что позволяет задерживать соль без потери калия.Напротив, при гиперкалиемии секреция калия благоприятна в дистальных отделах нефрона без значительного изменения скорости реабсорбции соли. Таким образом, калий выводится без удержания солей. То, как почки способны осуществлять две разные физиологические реакции в двух разных ситуациях (гиповолемия и гиперкалиемия), при которых повышен уровень альдостерона, известно как парадокс альдостерона [12]. Как почка распознает разницу между этими двумя состояниями? Как мы обсудим ниже, кажется вероятным, что реакция почек на эти два состояния объясняется наличием или отсутствием ангиотензина II (Ang II), уровень которого повышен при гиповолемии, но не при гиперкалиемии, и его регулирующим влиянием на транспорт ионов. пути через WNK4 в дистальном отделе нефрона.
То, как почки способны осуществлять две разные физиологические реакции в двух разных ситуациях (гиповолемия и гиперкалиемия), при которых повышен уровень альдостерона, известно как парадокс альдостерона [12]. Как почка распознает разницу между этими двумя состояниями? Как мы обсудим ниже, кажется вероятным, что реакция почек на эти два состояния объясняется наличием или отсутствием ангиотензина II (Ang II), уровень которого повышен при гиповолемии, но не при гиперкалиемии, и его регулирующим влиянием на транспорт ионов. пути через WNK4 в дистальном отделе нефрона.
Дистальный нефрон определяется как часть канальца, расположенная за пределами плотного пятна [13]. Изменения скорости транспорта соли в дистальном отделе нефрона больше не могут компенсироваться механизмом тубулогломерулярной обратной связи и, таким образом, оказывают существенное влияние на экскрецию соли с мочой. Дистальный отдел нефрона состоит из DCT, соединительных канальцев (CNT) и собирательных трубочек (CD). CD получает жидкость из нескольких сегментов DCT/CNT. Таким образом, в почках намного больше DCT/CNT, чем CD.DCT можно разделить на раннюю (DCT1) и позднюю (DCT2) части на основе экспрессии белка и гистологических различий [13,14,15]. Пути транспорта натрия в дистальном отделе нефрона включают NCC, который является мишенью для диуретиков тиазидного типа, экспрессируется исключительно в DCT [14,16], и эпителиальный натриевый канал (ENaC), мишень для амилорида, который экспрессируется в DCT2. и вдоль УНТ и CD [14,15,17]. Транспортные пути K + включают почечные наружные медуллярные калиевые каналы, ROMK и зависимые от потока большие Ca 2+ -активируемые калиевые каналы (BK каналы), оба из которых экспрессируются в дистальных отделах нефрона [18,19]. ,20].WNK1 и WNK4 охватывают весь дистальный отдел нефрона. Однако существует короткая специфичная для почек изоформа WNK1 (KS-WNK1), в которой отсутствует киназный домен, который стратегически экспрессируется только в DCT (см. ниже) [3,14,21,22]. Кроме того, было показано, что другая группа серин/треониновых киназ, известная как STE20-родственная пролин-аланин-богатая киназа SPAK и элемент окислительного стресса 1, OSR1, расположена ниже WNK [23,24].
Таким образом, в почках намного больше DCT/CNT, чем CD.DCT можно разделить на раннюю (DCT1) и позднюю (DCT2) части на основе экспрессии белка и гистологических различий [13,14,15]. Пути транспорта натрия в дистальном отделе нефрона включают NCC, который является мишенью для диуретиков тиазидного типа, экспрессируется исключительно в DCT [14,16], и эпителиальный натриевый канал (ENaC), мишень для амилорида, который экспрессируется в DCT2. и вдоль УНТ и CD [14,15,17]. Транспортные пути K + включают почечные наружные медуллярные калиевые каналы, ROMK и зависимые от потока большие Ca 2+ -активируемые калиевые каналы (BK каналы), оба из которых экспрессируются в дистальных отделах нефрона [18,19]. ,20].WNK1 и WNK4 охватывают весь дистальный отдел нефрона. Однако существует короткая специфичная для почек изоформа WNK1 (KS-WNK1), в которой отсутствует киназный домен, который стратегически экспрессируется только в DCT (см. ниже) [3,14,21,22]. Кроме того, было показано, что другая группа серин/треониновых киназ, известная как STE20-родственная пролин-аланин-богатая киназа SPAK и элемент окислительного стресса 1, OSR1, расположена ниже WNK [23,24]. SPAK и OSR1 присутствуют в толстом восходящем колене Генле, а также в DCT [25], а активация SPAK/OSR1 с помощью WNK модулирует взаимодействие/фосфорилирование этих киназ с переносчиками-мишенями.
SPAK и OSR1 присутствуют в толстом восходящем колене Генле, а также в DCT [25], а активация SPAK/OSR1 с помощью WNK модулирует взаимодействие/фосфорилирование этих киназ с переносчиками-мишенями.
Благодаря этим паттернам экспрессии существует четкая и понятная взаимосвязь между реабсорбцией натрия и секрецией калия в дистальном отделе нефрона. За пределами DCT, в CNT/CD, секреции калия способствуют повышенный кровоток и реабсорбция натрия (рис. 1). Это связано с тем, что вход Na + через ENaC генерирует отрицательное напряжение в просвете, что способствует секреции K + через ROMK [19]. Таким образом, для увеличения секреции калия необходимо увеличить доставку натрия к УНТ/ЦД.Это достигается за счет ингибирования вышестоящего переносчика реабсорбции соли NCC на ранней стадии DCT [26]. Кроме того, повышенная доставка жидкости к CNT/CD также активирует BK-каналы [27]. Именно по этим причинам увеличение поступления соли и объема к УНТ/ЦД петлевыми или тиазидными диуретиками (К + истощение) увеличивает экскрецию калия с мочой, в то время как К + -сберегающие диуретики, такие как амилорид, которые блокируют ENaC, косвенно предотвратить секрецию K + [6,28,29]. Эта взаимосвязь обработки дистальных отделов нефрона натрием и калием также объясняет положительный эффект исследования диетических подходов к остановке гипертензии (DASH) [30,31], в котором наилучшей комбинацией для предотвращения или улучшения гипертензии является диета с низким содержанием соли и высоким содержанием калия. , потому что для выведения избытка калия необходимо уменьшить реабсорбцию соли в DCT, тем самым увеличивая выведение соли почками [32].Это точное явление наблюдалось группой Палмера при изучении на крысах эффектов диеты с низким содержанием Na + с нормальным K + по сравнению с диетой с низким содержанием Na + с низким содержанием K + . В группе с низким содержанием K + экспрессия белка NCC резко увеличилась, что уменьшило дистальную доставку соли и Na + и ингибировало секрецию K + [33].
Эта взаимосвязь обработки дистальных отделов нефрона натрием и калием также объясняет положительный эффект исследования диетических подходов к остановке гипертензии (DASH) [30,31], в котором наилучшей комбинацией для предотвращения или улучшения гипертензии является диета с низким содержанием соли и высоким содержанием калия. , потому что для выведения избытка калия необходимо уменьшить реабсорбцию соли в DCT, тем самым увеличивая выведение соли почками [32].Это точное явление наблюдалось группой Палмера при изучении на крысах эффектов диеты с низким содержанием Na + с нормальным K + по сравнению с диетой с низким содержанием Na + с низким содержанием K + . В группе с низким содержанием K + экспрессия белка NCC резко увеличилась, что уменьшило дистальную доставку соли и Na + и ингибировало секрецию K + [33].
Рис. 1
a Во время гиповолемии увеличение Ang II изменяет активность WNK4, так что он стимулирует NCC и ENaC, ингибируя ROMK. Стимуляция NCC в DCT1 значительно снижает поток мочи вдоль дистальных отделов нефрона, тем самым сводя к минимуму секрецию K + . b Однако во время гиперкалиемии присутствие альдостерона в отсутствие Ang II приводит к ингибированию NCC в нечувствительном к альдостерону DCT1 и увеличению ENaC и ROMK из чувствительного к альдостерону DCT2 и далее. Снижение реабсорбции соли в DCT1 увеличивает приток мочи к CNT/CD, способствуя экскреции K + .
Стимуляция NCC в DCT1 значительно снижает поток мочи вдоль дистальных отделов нефрона, тем самым сводя к минимуму секрецию K + . b Однако во время гиперкалиемии присутствие альдостерона в отсутствие Ang II приводит к ингибированию NCC в нечувствительном к альдостерону DCT1 и увеличению ENaC и ROMK из чувствительного к альдостерону DCT2 и далее. Снижение реабсорбции соли в DCT1 увеличивает приток мочи к CNT/CD, способствуя экскреции K + .
Во время нормоволемии WNK4 предположительно ведет себя как ингибитор NCC, ENaC и ROMK [11,34,35]. Мутации PHA II изменяют способ, которым WNK4 регулирует эти транспортные системы. NCC и ENaC больше не ингибируются и, таким образом, предположительно активируются, в то время как ингибирование ROMK дополнительно усиливается мутантным WNK4 [11,34,35,36]. Это увеличивает реабсорбцию соли при минимизации секреции K + , что объясняет гипертензию и гиперкалиемию, которые характеризуют фенотип PHA II.Это крайняя, патофизиологическая версия того, к чему должна привести регулируемая реакция на низкий объем крови. Другими словами, WNK4, несущий мутации PHA II, имитирует нейрогормональное состояние, необходимое для сохранения объема перед лицом истощения внутрисосудистого объема, при котором желательным ответом является сохранение соли и объема без потери K + . Недавние данные подтверждают, что Ang II является активатором NCC [37,38,39], и наблюдение, что Ang II переключает поведение WNK4 с ингибитора на активатор NCC [40] и дополнительно усиливает ингибирование ROMK с помощью WNK4 [41]. ] предполагает, что мутации PHAII в WNK4 относятся к типу «усиления функции», имитируя эффект, который Ang II оказывает на NCC и ROMK.Таким образом, согласно этой модели, PHAII с мутациями в WNK4 может рассматриваться как следствие состояния «гиперангиотензинизма», локализованного в дистальном отделе нефрона, не потому, что Ang II повышен, а потому, что DCT/CNT работает так, как если бы Ang II был повышен. постоянно повышен.
Другими словами, WNK4, несущий мутации PHA II, имитирует нейрогормональное состояние, необходимое для сохранения объема перед лицом истощения внутрисосудистого объема, при котором желательным ответом является сохранение соли и объема без потери K + . Недавние данные подтверждают, что Ang II является активатором NCC [37,38,39], и наблюдение, что Ang II переключает поведение WNK4 с ингибитора на активатор NCC [40] и дополнительно усиливает ингибирование ROMK с помощью WNK4 [41]. ] предполагает, что мутации PHAII в WNK4 относятся к типу «усиления функции», имитируя эффект, который Ang II оказывает на NCC и ROMK.Таким образом, согласно этой модели, PHAII с мутациями в WNK4 может рассматриваться как следствие состояния «гиперангиотензинизма», локализованного в дистальном отделе нефрона, не потому, что Ang II повышен, а потому, что DCT/CNT работает так, как если бы Ang II был повышен. постоянно повышен.
У пациентов с PHA II из-за сверхэкспрессии WNK1 объяснение активации NCC и ингибирования ROMK следующее: во-первых, WNK1 напрямую ингибирует ROMK [42,43,44,45,46]. Следовательно, любое увеличение экспрессии WNK1 приводит к дозозависимому снижению ROMK.Кроме того, WNK1 является ингибитором WNK4, но этот эффект, в свою очередь, предотвращается KS-WNK1, короткой изоформой, в которой отсутствует киназный домен и которая экспрессируется только в DCT (рис. 2). В физиологических условиях KS-WNK1 притупляет действие WNK1 на WNK4 и, таким образом, NCC свободно ингибируется WNK4 [47,48]. Однако у пациентов с PHA II усиленная экспрессия WNK1 увеличивает соотношение WNK1/KS-WNK1, преодолевая ингибирующий эффект KS-WNK1, что позволяет WNK1 ингибировать WNK4, что приводит к индуцированной WNK1 активации NCC (рис.2). Таким образом, гипертензия у пациентов WNK1 PHAII, по-видимому, также является следствием повышенной активности NCC в DCT, аналогично тому, что происходит у пациентов WNK4 PHA II и во время гиповолемии.
Следовательно, любое увеличение экспрессии WNK1 приводит к дозозависимому снижению ROMK.Кроме того, WNK1 является ингибитором WNK4, но этот эффект, в свою очередь, предотвращается KS-WNK1, короткой изоформой, в которой отсутствует киназный домен и которая экспрессируется только в DCT (рис. 2). В физиологических условиях KS-WNK1 притупляет действие WNK1 на WNK4 и, таким образом, NCC свободно ингибируется WNK4 [47,48]. Однако у пациентов с PHA II усиленная экспрессия WNK1 увеличивает соотношение WNK1/KS-WNK1, преодолевая ингибирующий эффект KS-WNK1, что позволяет WNK1 ингибировать WNK4, что приводит к индуцированной WNK1 активации NCC (рис.2). Таким образом, гипертензия у пациентов WNK1 PHAII, по-видимому, также является следствием повышенной активности NCC в DCT, аналогично тому, что происходит у пациентов WNK4 PHA II и во время гиповолемии.
Рис. 2
a В физиологических условиях в DCT1 экспрессия короткой изоформы KS-WNK1 в несколько раз выше, чем длинного варианта WNK1. Таким образом, KS-WNK1 предотвращает индуцированное WNK1 ингибирование WNK4 и, таким образом, позволяет WNK4 свободно ингибировать NCC и ROMK. b Напротив, в PHAII из-за интронных делеций WNK1 повышенная экспрессия варианта long-WNK1 преодолевает ингибирование KS-WNK1, приводящее к активации NCC, и ингибирование ROMK как следствие ингибирования WNK4 WNK1.Кроме того, длинный WNK1 напрямую ингибирует ROMK.
Таким образом, KS-WNK1 предотвращает индуцированное WNK1 ингибирование WNK4 и, таким образом, позволяет WNK4 свободно ингибировать NCC и ROMK. b Напротив, в PHAII из-за интронных делеций WNK1 повышенная экспрессия варианта long-WNK1 преодолевает ингибирование KS-WNK1, приводящее к активации NCC, и ингибирование ROMK как следствие ингибирования WNK4 WNK1.Кроме того, длинный WNK1 напрямую ингибирует ROMK.
Другим членом семейства киназ WNK, который сам по себе также может имитировать это нейрогормональное состояние задержки соли без экскреции калия, является WNK3. Эта киназа, которая экспрессируется в DCT и CCD, активирует NCC [49], а также ингибирует ROMK [50]. Взаимодействие/конкуренция между WNK3 и WNK4 в отношении NCC является еще одним потенциальным механизмом модуляции скорости транспорта соли при DCT [51].
Эти довольно сложные механизмы регуляции транспорта начинают объяснять наблюдаемые сложные клинические проявления.На самом деле, если посмотреть на общие механизмы, реакция почек на различные заболевания становится очевидной (таблица 1). Заболевания, которые проявляются низким объемом крови, будь то прямая кровопотеря или аномалии распределения внеклеточной жидкости, такие как хроническая сердечная недостаточность, цирроз печени, нефротический синдром и другие, имеют по существу одну и ту же основную проблему: снижение среднего циркуляторного давления наполнения и снижение потока мочи в дистальный отдел нефрона. У этих больных активируется ренин-ангиотензин-альдостероновая система (вторичный альдостеронизм).Благодаря механизмам, описанным выше (рис. 1), при вторичном альдостеронизме снижается кровоток в дистальном отделе нефрона, что приводит к состоянию удержания объема, которое пытается увеличить объем циркулирующей крови без изменения K + плазмы. Снижение потока мочи в этой части нефрона способствует задержке K + за счет ингибирования каналов BK и косвенного ингибирования механизма обмена ENaC/ROMK Na + /K + . Таким образом, у нелеченых пациентов с состояниями, при которых происходит активация как Ang II, так и альдостерона, наблюдается индуцированная Ang II-WNK4 и альдостероном задержка Na + и объема из-за активации NCC и ENaC, но без изменений в уровень K в плазме + , потому что индуцированное альдостероном увеличение экспрессии/активности ROMK преодолевается снижением активности ROMK Ang II-WNK4 и WNK1, а также снижением потока и доставки соли к CNT/CD из-за увеличения реабсорбция солей при ДСТ.
Заболевания, которые проявляются низким объемом крови, будь то прямая кровопотеря или аномалии распределения внеклеточной жидкости, такие как хроническая сердечная недостаточность, цирроз печени, нефротический синдром и другие, имеют по существу одну и ту же основную проблему: снижение среднего циркуляторного давления наполнения и снижение потока мочи в дистальный отдел нефрона. У этих больных активируется ренин-ангиотензин-альдостероновая система (вторичный альдостеронизм).Благодаря механизмам, описанным выше (рис. 1), при вторичном альдостеронизме снижается кровоток в дистальном отделе нефрона, что приводит к состоянию удержания объема, которое пытается увеличить объем циркулирующей крови без изменения K + плазмы. Снижение потока мочи в этой части нефрона способствует задержке K + за счет ингибирования каналов BK и косвенного ингибирования механизма обмена ENaC/ROMK Na + /K + . Таким образом, у нелеченых пациентов с состояниями, при которых происходит активация как Ang II, так и альдостерона, наблюдается индуцированная Ang II-WNK4 и альдостероном задержка Na + и объема из-за активации NCC и ENaC, но без изменений в уровень K в плазме + , потому что индуцированное альдостероном увеличение экспрессии/активности ROMK преодолевается снижением активности ROMK Ang II-WNK4 и WNK1, а также снижением потока и доставки соли к CNT/CD из-за увеличения реабсорбция солей при ДСТ. Поскольку увеличение объема циркулирующей крови происходит за счет увеличения Na + и воды, что не сопровождается сопутствующим повышением онкотического давления, это приводит к накоплению жидкости во всем межклеточном пространстве, что клинически будет проявляться отеком (табл. 1).
Поскольку увеличение объема циркулирующей крови происходит за счет увеличения Na + и воды, что не сопровождается сопутствующим повышением онкотического давления, это приводит к накоплению жидкости во всем межклеточном пространстве, что клинически будет проявляться отеком (табл. 1).
Таблица 1
Общие клинические проявления и связанные с ними изменения альдостерона, Ang II, артериального давления, баланса Na+ и баланса K+
Совершенно другая ситуация возникает, когда альдостерон повышается при отсутствии активации ренин-ангиотензин-альдостероновой системы. система (рис.1б). Это то, что происходит во время диеты с высоким содержанием калия или при первичном альдостеронизме, при котором увеличивается скорость потока в дистальных отделах нефрона. Альдостерон увеличивает экспрессию сывороточной глюкокортикоидкиназы 1 (SGK1), которая, в свою очередь, фосфорилирует и ингибирует белок, известный как Nedd4-2, который путем убиквитилирования ENaC запускает интернализацию и разрушение канала. Таким образом, ингибируя Nedd4-2, SGK1 индуцирует и увеличивает экспрессию ENaC в плазматической мембране [52]. Кроме того, путем фосфорилирования WNK4 в двух критических сайтах, SGK1 также будет предотвращать индуцированное WNK4 ингибирование ENaC и ROMK, тем самым высвобождая активность этих каналов, что способствует секреции K + через механизм обмена ENaC/ROMK. см. выше) [35,53].Также известно, что альдостерон увеличивает экспрессию белка NCC [54] посредством регуляции Nedd4-2, механизм, аналогичный механизму ENaC [55]. Однако опосредованное альдостероном увеличение NCC, по-видимому, будет ограничено DCT2, где количество NCC относительно невелико, потому что DCT1 не рассматривается как часть чувствительного к альдостерону дистального отдела нефрона из-за отсутствия 11β-гидроксистероиддегидрогеназы 2 типа, что исключает беспорядочное занятие минералокортикоидных рецепторов кортизолом [22,56].Следовательно, хотя экспрессия NCC увеличивается в DCT2 в состоянии изолированной секреции альдостерона, из-за отсутствия Ang II, NCC в DCT1 остается при ингибировании WNK4.
Таким образом, ингибируя Nedd4-2, SGK1 индуцирует и увеличивает экспрессию ENaC в плазматической мембране [52]. Кроме того, путем фосфорилирования WNK4 в двух критических сайтах, SGK1 также будет предотвращать индуцированное WNK4 ингибирование ENaC и ROMK, тем самым высвобождая активность этих каналов, что способствует секреции K + через механизм обмена ENaC/ROMK. см. выше) [35,53].Также известно, что альдостерон увеличивает экспрессию белка NCC [54] посредством регуляции Nedd4-2, механизм, аналогичный механизму ENaC [55]. Однако опосредованное альдостероном увеличение NCC, по-видимому, будет ограничено DCT2, где количество NCC относительно невелико, потому что DCT1 не рассматривается как часть чувствительного к альдостерону дистального отдела нефрона из-за отсутствия 11β-гидроксистероиддегидрогеназы 2 типа, что исключает беспорядочное занятие минералокортикоидных рецепторов кортизолом [22,56].Следовательно, хотя экспрессия NCC увеличивается в DCT2 в состоянии изолированной секреции альдостерона, из-за отсутствия Ang II, NCC в DCT1 остается при ингибировании WNK4. Это приводит к увеличению дистальной доставки Na + и увеличению объема и активации каналов ENaC, ROMK и BK, что приводит к увеличению обмена Na + /K + и, таким образом, увеличению экскреции K + . Это явление становится клинически значимым при состояниях повышенной секреции альдостерона со сниженной активностью Ang II и, как следствие, усилением кровотока в дистальных отделах нефрона (таблица 1).Повседневный сценарий, который имитирует эту ситуацию, — это диета с высоким содержанием K + , при которой реабсорбция соли в DCT1 снижается, что позволяет обмену Na + / K + в CD секретировать избыток K + . Патофизиологическим вариантом является первичный альдостеронизм, при котором автономное повышение уровня альдостерона способствует реабсорбции натрия и секреции К + за счет повышения активности ENaC и ROMK соответственно. Почему у этих пациентов не развивается отек, подобно тому, что происходит у пациентов с вторичным гиперальдостеронизмом, связано с наличием или отсутствием Ang II при вторичном или первичном альдостеронизме.
Это приводит к увеличению дистальной доставки Na + и увеличению объема и активации каналов ENaC, ROMK и BK, что приводит к увеличению обмена Na + /K + и, таким образом, увеличению экскреции K + . Это явление становится клинически значимым при состояниях повышенной секреции альдостерона со сниженной активностью Ang II и, как следствие, усилением кровотока в дистальных отделах нефрона (таблица 1).Повседневный сценарий, который имитирует эту ситуацию, — это диета с высоким содержанием K + , при которой реабсорбция соли в DCT1 снижается, что позволяет обмену Na + / K + в CD секретировать избыток K + . Патофизиологическим вариантом является первичный альдостеронизм, при котором автономное повышение уровня альдостерона способствует реабсорбции натрия и секреции К + за счет повышения активности ENaC и ROMK соответственно. Почему у этих пациентов не развивается отек, подобно тому, что происходит у пациентов с вторичным гиперальдостеронизмом, связано с наличием или отсутствием Ang II при вторичном или первичном альдостеронизме. При вторичном альдостеронизме, как обсуждалось выше, Ang II, предположительно через WNK4, активирует NCC. Напротив, при первичном альдостеронизме тот факт, что NCC не активируется альдостероном при DCT1, свидетельствует о том, что эти пациенты все еще подвержены натрийурезу под давлением. На самом деле, хотя NCC считается белком, регулируемым альдостероном [54], во время феномена ускользания альдостерона из всех исследованных транспортеров в почках NCC является единственным, чья экспрессия действительно снижена, несмотря на постоянное введение альдостерон [57].Таким образом, при первичном альдостеронизме сниженная активность NCC в DCT1 поддерживает солевой баланс и за счет увеличения скорости потока обеспечивает доставку соли и объема к CNT/CD, вызывая гипокалиемию. Иными словами, ингибирования NCC в DCT1 достаточно, чтобы преодолеть активность ENaC хотя бы для предотвращения образования отека, но за счет артериальной гипертензии. Этот механизм распространяется на синдром Кушинга. Когда повышенный уровень кортизола в плазме превышает способность 11β-гидроксистероиддегидрогеназы 2 типа разрушать его и, таким образом, занимает и активирует минералокортикоидный рецептор, как если бы уровень альдостерона был повышен.
При вторичном альдостеронизме, как обсуждалось выше, Ang II, предположительно через WNK4, активирует NCC. Напротив, при первичном альдостеронизме тот факт, что NCC не активируется альдостероном при DCT1, свидетельствует о том, что эти пациенты все еще подвержены натрийурезу под давлением. На самом деле, хотя NCC считается белком, регулируемым альдостероном [54], во время феномена ускользания альдостерона из всех исследованных транспортеров в почках NCC является единственным, чья экспрессия действительно снижена, несмотря на постоянное введение альдостерон [57].Таким образом, при первичном альдостеронизме сниженная активность NCC в DCT1 поддерживает солевой баланс и за счет увеличения скорости потока обеспечивает доставку соли и объема к CNT/CD, вызывая гипокалиемию. Иными словами, ингибирования NCC в DCT1 достаточно, чтобы преодолеть активность ENaC хотя бы для предотвращения образования отека, но за счет артериальной гипертензии. Этот механизм распространяется на синдром Кушинга. Когда повышенный уровень кортизола в плазме превышает способность 11β-гидроксистероиддегидрогеназы 2 типа разрушать его и, таким образом, занимает и активирует минералокортикоидный рецептор, как если бы уровень альдостерона был повышен. Аналогичная ситуация возникает при синдроме Лиддла, при котором происходит первичное повышение активности ENaC из-за мутаций в β- или γ-субъединицах ENaC, которые устраняют мотив PY, необходимый для взаимодействия Nedd4–2 и ENaC, что препятствует нормальной регуляции канала. по Недду4–2 [58]. Клиническая картина синдрома Лиддла сходна с первичным альдостеронизмом: гипертензия с гипокалиемией. Повышенная абсорбция натрия ENaC вызывает гипертензию, а механизмы натрийуреза под давлением должны ингибировать NCC для поддержания солевого баланса, предотвращая образование отеков.
Аналогичная ситуация возникает при синдроме Лиддла, при котором происходит первичное повышение активности ENaC из-за мутаций в β- или γ-субъединицах ENaC, которые устраняют мотив PY, необходимый для взаимодействия Nedd4–2 и ENaC, что препятствует нормальной регуляции канала. по Недду4–2 [58]. Клиническая картина синдрома Лиддла сходна с первичным альдостеронизмом: гипертензия с гипокалиемией. Повышенная абсорбция натрия ENaC вызывает гипертензию, а механизмы натрийуреза под давлением должны ингибировать NCC для поддержания солевого баланса, предотвращая образование отеков.
Таким образом, интеграция почечных молекулярных механизмов, которые регулируются с помощью киназ WNK, пролила свет на проблему, которая долгое время оставалась неясной. Молекулярные механизмы взаимодействия между альдостероном и Ang II в дифференциации между задержкой объема и секрецией K + начинают проясняться, обеспечивая лучшее молекулярное понимание физиологических или патофизиологических процессов, связанных с классическими клиническими синдромами солевого и калиевого обмена.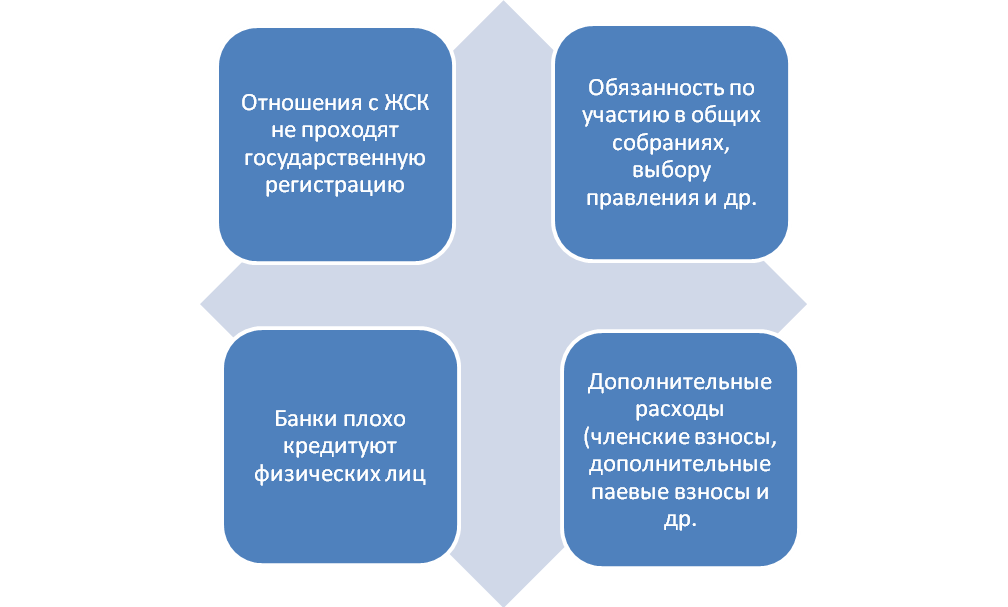
Благодарности
Мы благодарим Пола Киммела, доктора медицины, за его критическое прочтение рукописи. Эта работа была частично поддержана Трансатлантической сетью Фонда Leducq по гипертензии и грантом 165815 El Consejo Nacional de Ciencia y Tecnología (CONACYT-Mexico) (GG). J.P.A. получил стипендию от CONACYT-Mexico и является аспирантом программы PhD в области биомедицинских наук Национального автономного университета Мексики, UNAM.
Заявление о раскрытии информации
Конфликт интересов не заявлен
Ссылки
- Хорн Э.Дж., Нельсон, Дж.Х., Маккормик, Дж.А., Эллисон, Д.Х.: Сеть киназ WNK, регулирующая натрий, калий и артериальное давление.J Am Soc Nephrol 2011;22:605–614.
- Маккормик Дж. А., Эллисон Д. Х.: WNK: атипичные протеинкиназы с плейотропным действием. Physiol Rev 2011; 91: 177–219.
- Уилсон Ф.Х., Диссе-Никодем С., Чоат К.А., Исикава К., Нельсон-Уильямс С., Дезиттер И., Гюнель М., Милфорд Д.В., Липкин Г.В., Ачард Дж.М., Фили М.П., Дюссол Б., Берланд И., Анвин Р.Дж., Майан Х., Саймон DB, Farfel Z, Jeunemaitre X, Lifton RP: Гипертония человека, вызванная мутациями в киназах WNK.Наука 2001; 293:1107–1112.
- Mayan H, Vered I, Mouallem M, Tzadok-Witkon M, Pauzner R, Farfel Z: Псевдогипоальдостеронизм II типа: выраженная чувствительность к тиазидам, гиперкальциурия, нормомагниемия и низкая минеральная плотность костей. J Clin Endocrinol Metab 2002;87:3248–3254.
- Гордон Р.Д., Ходсман Г.П. Синдром гипертензии и гиперкалиемии без почечной недостаточности: длительная коррекция тиазидным диуретиком.Скотт Мед Дж. 1986; 31:43–44.
- Gamba G: котранспортер Na + -Cl – , чувствительный к тиазидам: молекулярная биология, функциональные свойства и регуляция с помощью WNK. Am J Physiol Renal Physiol 2009; 297: F838–F848.
- Саймон Д. Б., Нельсон-Уильямс С., Джонсон-Биа М., Эллисон Д., Карет Ф.Е., Морей-Молина А., Ваара И., Ивата Ф., Кушнер Х.М., Кулен М., Гаинза Ф.Дж., Гительман Х.Дж., Лифтон Р.П.: Гительмановский вариант синдрома Барттера наследственный гипокалиемический алкалоз вызывается мутациями в тиазид-чувствительном котранспортере Na-Cl.Нат Жене 1996; 12:24–30.
- Плоткин М.Д., Каплан М.Р., Верландер Дж.М., Ли В.С., Браун Д., Поч Э., Гулланс С.Р., Хеберт С.К.: Локализация чувствительного к тиазиду котранспортера Na-Cl, rTSC1, в почках крыс. Kidney Int 1996; 50: 174–183.
- Loffing J, Vallon V, Loffing-Cueni D, Aregger F, Richter K, Pietri L, Bloch-Faure M, Hoenderop JG, Shull GE, Meneton P, Kaissling B: измененная структура дистальных канальцев почек и почечный Na + и Ca 2+ обращение с мышами в модели синдрома Гительмана. J Am Soc Nephrol 2004;15:2276–2288.
- Бахманн С., Веласкес Х., Обермюллер Н., Рейли Р.Ф., Мозер Д., Эллисон Д.Х.: Экспрессия тиазид-чувствительного котранспортера Na-Cl клетками дистальных извитых канальцев кролика. Дж. Клин инвест 1995; 96: 2510–2514.
- Wilson FH, Kahle KT, Sabath E, Lalioti MD, Rapson AK, Hoover RS, Hebert SC, Gamba G, Lifton RP: Молекулярный патогенез наследственной гипертензии с гиперкалиемией: котранспортер Na-Cl ингибируется диким типом, но не мутантным WNK4 .Proc Natl Acad Sci USA 2003;100:680–684.
- Arroyo JP, Ronzaud C, Lagnaz D, Staub O, Gamba G: Парадокс альдостерона: дифференциальная регуляция транспорта ионов в дистальных отделах нефрона. Физиология (Bethesda) 2011; 26: 115–123.
- Рейли Р.Ф., Эллисон Д.Х.: Дистальные канальцы млекопитающих: физиология, патофизиология и молекулярная анатомия.Physiol Rev 2000; 80: 277–313.
- Loffing J, Loffing-Cueni D, Valderrabano V, Klausli L, Hebert SC, Rossier BC, Hoenderop JG, Bindels RJ, Kaissling B: Распределение путей трансцеллюлярного транспорта кальция и натрия вдоль дистальных отделов нефрона мыши. Am J Physiol Renal Physiol 2001; 281: F1021–F1027.
- Бинер Х. Л., Арпин-Ботт М.П., Лоффинг Дж., Ван Х, Неппер М., Хеберт С.К., Кайслинг Б.: Кортикальный дистальный нефрон человека: распределение путей транспорта электролита и воды. J Am Soc Nephrol 2002;13:836–847.
- Эллисон Д.Х., Веласкес Х., Райт Ф.С.: Котранспорт хлорида натрия, чувствительный к тиазидам, в ранних дистальных канальцах.Am J Physiol Renal Fluid Electrolyte Physiol 1987;253:F546–F554.
- Лоффинг Дж., Кайсслинг Б.: Пути транспорта натрия и кальция вдоль дистального отдела нефрона млекопитающих: от кролика к человеку. Am J Physiol Renal Physiol 2003; 284: F628–F643.
- Аморим Дж. Б., Бейли М. А., Муса-Азиз Р., Гибиш Г., Малник Г.: Роль просветного аниона и рН в секреции калия дистальными канальцами.Am J Physiol Renal Physiol 2003; 284: F381–F388.
- Хеберт С.К., Дезир Г., Гибиш Г., Ван В.: Молекулярное разнообразие и регуляция почечных калиевых каналов. Physiol Rev 2005; 85: 319–371.
- Grimm PR, Sansom SC: каналы BK и новая форма гипертонии.Kidney Int 2010; 78: 956–962.
- О’Рейли М. , Маршалл Э., Макгилливрей Т., Миттал М., Сюэ В., Кеньон С.Дж., Браун Р.В.: Реакции, обусловленные диетическими электролитами, в почечном пути киназы WNK in vivo. J Am Soc Nephrol 2006;17:2402–2413.
- Бахманн С., Бостанджогло М., Шмитт Р., Эллисон Д.Х.: Белки, связанные с транспортом натрия, в дистальном нефроне млекопитающих – распределение, онтогенез и функциональные аспекты.Анат Эмбриол (Берл) 1999; 200:447–468.
- Пьехотта К., Лу Дж., Дельпир Э.: Котранспортеры хлоридов катионов взаимодействуют со связанными со стрессом киназами Ste20-родственной пролин-аланин-богатой киназой (SPAK) и реакцией на окислительный стресс 1 (OSR1). J Biol Chem 2002; 277:50812–50819.
- Vitari AC, Deak M, Morrice NA, Alessi DR: Протеинкиназы WNK1 и WNK4, которые мутируют при синдроме гипертонии Гордона, фосфорилируют и активируют протеинкиназы SPAK и OSR1.Биохим Дж. 2005; 391:17–24.
- Рафики Ф.Х., Зубер А.М., Гловер М., Ричардсон С., Флеминг С., Йованович С., Йованович А., О’Шонесси К.М., Алесси Д.Р.: Роль активируемой WNK киназы SPAK в регулировании артериального давления. EMBO Mol Med 2010; 2: 63–75.
- Фриндт Г. , Палмер Л.Г.: Влияние диетического калия на экспрессию почечных ионных каналов и транспортеров на клеточной поверхности.Am J Physiol Renal Physiol 2010;299:F890–F897.
- Вода С.Б., Брагин А., Клейман Т.Р., Сатлин Л.М.: Зависимая от потока секреция К + в кортикальных собирательных трубочках опосредована макси-К каналом. Am J Physiol Renal Physiol 2001;280:F786–F793.
- Кришна Г.Г., Шульман М.Д., Наринс Р.Г. Клиническое применение калийсберегающих диуретиков.Семин Нефрол 1988; 8: 354–364.
- Роуз Б. Д.: Мочегонные средства. Kidney Int 1991; 39: 336–352.
- Appel LJ, Moore TJ, Obarzanek E, Vollmer WM, Svetkey LP, Sacks FM, Bray GA, Vogt TM, Cutler JA, Windhauser MM, Lin PH, Karanja N: Клиническое исследование влияния режима питания на кровяное давление.Совместная исследовательская группа DASH. N Engl J Med 1997; 336:1117–1124.
- Сакс Ф.М., Светкей Л.П., Воллмер В.М., Аппель Л.Дж., Брей Г.А., Харша Д., Обаржанек Э., Конлин П.Р., Миллер Э.Р. III, Саймонс-Мортон Д.Г., Каранджа Н., Лин П.Х.: Влияние на артериальное давление пониженного содержания натрия в рационе и рациона питания Подходы к диете Stop Hypertension (DASH). Совместная исследовательская группа DASH-Sodium. N Engl J Med 2001; 344: 3–10.
- Huang CL, Kuo E: Механизмы заболевания: WNK-ing в механизме солечувствительной гипертензии. Nat Clin Pract Nephrol 2007; 3: 623–630.
- Frindt G, Houde V, Palmer LG: Сохранение Na + vs.К + по корковому отделу собирательных трубочек крысы. Am J Physiol Renal Physiol 2011;301:F14–F20.
- Kahle KT, Wilson FH, Leng Q, Lalioti MD, O’Connell AD, Dong K, Rapson AK, MacGregor GG, Giebisch G, Hebert SC, Lifton RP: WNK4 регулирует баланс между реабсорбцией NaCl в почках и секрецией K + . Нат Жене 2003; 35: 372–376.
- Ring AM, Cheng SX, Leng Q, Kahle KT, Rinehart J, Lalioti MD, Volkman HM, Wilson FH, Hebert SC, Lifton RP: WNK4 регулирует активность эпителиального канала Na + in vitro и in vivo. Proc Natl Acad Sci USA 2007;104:4020–4024.
- Сан-Кристобаль П., Де Лос Х.П., Понсе-Кориа Дж., Морено Э., Гамба Г.: Киназы WNK, транспорт ионов в почках и гипертония.Am J Nephrol 2008; 28: 860–870.
- Сандберг М. Б., Рикье А.Д., Пихакаски-Маунсбах К., Макдоноу А.А., Маунсбах А.Б.: Ангиотензин II провоцирует острую транспортировку котранспортера NaCl (NCC) из дистальных канальцев к апикальной мембране. Am J Physiol Renal Physiol 2007; 293: F662–F669.
- van der LN, Lim CH, Fenton RA, Meima ME, Jan Danser AH, Zietse R, Hoorn EJ: Ангиотензин II индуцирует фосфорилирование тиазид-чувствительного котранспортера хлорида натрия независимо от альдостерона.Kidney Int 2011; 79: 66–76.
- Чжао Д., Сет Д.М., Навар Л.Г.: Повышенная реабсорбция натрия в дистальных отделах нефрона у мышей, которым постоянно вводили ангиотензин II. Гипертония 2009;54:120–126.
- San Cristobal P, Pacheco-Alvarez D, Richardson C, Ring AM, Vazquez N, Rafiqi FH, Chari D, Kahle KT, Leng Q, Bobadilla NA, Hebert SC, Alessi DR, Lifton RP, Gamba G: передача сигналов ангиотензина II увеличивает активность почечного котранспортера Na-Cl через WNK4-SPAK-зависимый путь.Proc Natl Acad Sci USA 2009;106:4384–4389.
- Yue P, Sun P, Lin DH, Pan C, Xing W, Wang W: Ангиотензин II уменьшает влияние SGK1 на опосредованное WNK4 ингибирование каналов ROMK1. Kidney Int 2011; 79: 423–431.
- Cheng CJ, Huang CL: Активация PI3-киназы стимулирует эндоцитоз ROMK посредством Akt1/SGK1-зависимого фосфорилирования WNK1. J Am Soc Nephrol 2011;22:460–471.
- He G, Wang HR, Huang SK, Huang CL: Intersectin связывает киназы WNK с эндоцитозом ROMK1. J Clin Invest 2007; 117: 1078–1087.
- Huang CL, Yang SS, Lin SH: Механизм регуляции почечного транспорта ионов киназами WNK.Curr Opin Nephrol Hypertens 2008;17:519–525.
- Лазрак А., Лю З., Хуанг С.Л.: Антагонистическая регуляция ROMK длинными и почечными изоформами WNK1. Proc Natl Acad Sci USA 2006;103:1615–1620.
- Liu Z, Wang HR, Huang CL: Регуляция канала ROMK и гомеостаза K + почечной киназой WNK1.J Biol Chem 2009; 284:12198–12206.
- Субраманья А.Р., Ян С.Л., Чжу С., Эллисон Д.Х.: Доминантно-негативная регуляция WNK1 его дефектной почечной киназой изоформой. Am J Physiol Renal Physiol 2006; 290: F619–F624.
- Liu Z, Xie J, Wu T, Truong T, Auchus RJ, Huang CL: Понижающая регуляция котранспортеров NCC и NKCC2 почечно-специфическим WNK1, выявленная с помощью разрушения генов и моделей трансгенных мышей. Hum Mol Genet 2011; 20: 855–866.
- Rinehart J, Kahle KT, De Los HP, Vazquez N, Meade P, Wilson FH, Hebert SC, Gimenez I, Gamba G, Lifton RP: киназа WNK3 является положительным регулятором NKCC2 и NCC, почечных котранспортеров катионов-Cl, необходимых для нормальной гомеостаз артериального давления.Proc Natl Acad Sci USA 2005;102:16777–16782.
- Leng Q, Kahle KT, Rinehart J, MacGregor GG, Wilson FH, Canessa CM, Lifton RP, Hebert SC: WNK3, киназа, связанная с генами, мутировавшими при наследственной гипертензии с гиперкалиемией, регулирует канал K + ROMK1 (Kir1.1). J Physiol 2006; 571: 275–286.
- Yang CL, Zhu X, Ellison DH: Котранспортер Na-Cl, чувствительный к тиазидам, регулируется сигнальным комплексом киназы WNK. J Clin Invest 2007;117:3403–3411.
- Абриэль Х., Штауб О.: Убиквитинирование ионных каналов.Физиология (Bethesda) 2005; 20: 398–407.
- Ring AM, Leng Q, Rinehart J, Wilson FH, Kahle KT, Hebert SC, Lifton RP: сайт SGK1 в WNK4 регулирует активность каналов Na + и каналов K + и имеет значение для передачи сигналов альдостерона и K + . гомеостаз.Proc Natl Acad Sci USA 2007;104:4025–4029.
- Ким Г.Х., Масиламани С., Тернер Р., Митчелл С., Уэйд Дж. Б., Неппер М.А.: Котранспортер Na-Cl, чувствительный к тиазидам, представляет собой белок, индуцированный альдостероном. Proc Natl Acad Sci USA 1998; 95:14552–14557.
- Arroyo JP, Lagnaz D, Ronzaud C, Vazquez N, Ko BS, Moddes L, Ruffieux-Dadidié D, Hausel P, Koesters R, Yang B, Stokes JB, Hoover RS, Gamba G, Staub O: Nedd4-2 модулирует почечный Na +– Cl – котранспортер по пути альдостерон-SGK1-Nedd4-2.J Am Soc Nephrol 2011; 22: 1707–1719.
- Бостанджогло М. , Ривз В.Б., Рейли Р.Ф., Веласкес Х., Робертсон Н., Литвак Г., Морсинг П., Доруп Дж., Бахманн С., Эллисон Д.Х., Бостонджогло М.: экспрессия 11-бета-гидроксистероиддегидрогеназы, минералокортикоидного рецептора и тиазид-чувствительного котранспортера Na-Cl дистальными канальцами.J Am Soc Nephrol 1998; 9: 1347–1358.
- Wang XY, Masilamani S, Nielsen J, Kwon TH, Brooks HL, Nielsen S, Knepper MA: Почечный тиазид-чувствительный котранспортер Na-Cl как медиатор феномена ускользания альдостерона. J Clin Invest 2001; 108: 215–222.
- Шимкетс Р.А., Варнок Д.Г., Боситис К.М., Нельсон-Уильямнс С. , Ханссон Дж.Х., Шамбелан М., Гилл Дж.Р., Улик С., Милора Р.В., Финдлинг Дж.В., Канесса С.М., Россье Б.К., Лифтон Р.П.: Синдром Лиддла: наследственная человеческая гипертензия, вызванная мутациями в бета-субъединице эпителиального натриевого канала.Ячейка 1994; 79: 407–414.
 А., Эллисон Д. Х.: WNK: атипичные протеинкиназы с плейотропным действием. Physiol Rev 2011; 91: 177–219.
А., Эллисон Д. Х.: WNK: атипичные протеинкиназы с плейотропным действием. Physiol Rev 2011; 91: 177–219. J Clin Endocrinol Metab 2002;87:3248–3254.
J Clin Endocrinol Metab 2002;87:3248–3254. Б., Нельсон-Уильямс С., Джонсон-Биа М., Эллисон Д., Карет Ф.Е., Морей-Молина А., Ваара И., Ивата Ф., Кушнер Х.М., Кулен М., Гаинза Ф.Дж., Гительман Х.Дж., Лифтон Р.П.: Гительмановский вариант синдрома Барттера наследственный гипокалиемический алкалоз вызывается мутациями в тиазид-чувствительном котранспортере Na-Cl.Нат Жене 1996; 12:24–30.
Б., Нельсон-Уильямс С., Джонсон-Биа М., Эллисон Д., Карет Ф.Е., Морей-Молина А., Ваара И., Ивата Ф., Кушнер Х.М., Кулен М., Гаинза Ф.Дж., Гительман Х.Дж., Лифтон Р.П.: Гительмановский вариант синдрома Барттера наследственный гипокалиемический алкалоз вызывается мутациями в тиазид-чувствительном котранспортере Na-Cl.Нат Жене 1996; 12:24–30. J Am Soc Nephrol 2004;15:2276–2288.
J Am Soc Nephrol 2004;15:2276–2288. Физиология (Bethesda) 2011; 26: 115–123.
Физиология (Bethesda) 2011; 26: 115–123. Л., Арпин-Ботт М.П., Лоффинг Дж., Ван Х, Неппер М., Хеберт С.К., Кайслинг Б.: Кортикальный дистальный нефрон человека: распределение путей транспорта электролита и воды. J Am Soc Nephrol 2002;13:836–847.
Л., Арпин-Ботт М.П., Лоффинг Дж., Ван Х, Неппер М., Хеберт С.К., Кайслинг Б.: Кортикальный дистальный нефрон человека: распределение путей транспорта электролита и воды. J Am Soc Nephrol 2002;13:836–847. Б., Бейли М. А., Муса-Азиз Р., Гибиш Г., Малник Г.: Роль просветного аниона и рН в секреции калия дистальными канальцами.Am J Physiol Renal Physiol 2003; 284: F381–F388.
Б., Бейли М. А., Муса-Азиз Р., Гибиш Г., Малник Г.: Роль просветного аниона и рН в секреции калия дистальными канальцами.Am J Physiol Renal Physiol 2003; 284: F381–F388. , Маршалл Э., Макгилливрей Т., Миттал М., Сюэ В., Кеньон С.Дж., Браун Р.В.: Реакции, обусловленные диетическими электролитами, в почечном пути киназы WNK in vivo. J Am Soc Nephrol 2006;17:2402–2413.
, Маршалл Э., Макгилливрей Т., Миттал М., Сюэ В., Кеньон С.Дж., Браун Р.В.: Реакции, обусловленные диетическими электролитами, в почечном пути киназы WNK in vivo. J Am Soc Nephrol 2006;17:2402–2413. J Biol Chem 2002; 277:50812–50819.
J Biol Chem 2002; 277:50812–50819. , Палмер Л.Г.: Влияние диетического калия на экспрессию почечных ионных каналов и транспортеров на клеточной поверхности.Am J Physiol Renal Physiol 2010;299:F890–F897.
, Палмер Л.Г.: Влияние диетического калия на экспрессию почечных ионных каналов и транспортеров на клеточной поверхности.Am J Physiol Renal Physiol 2010;299:F890–F897. Д.: Мочегонные средства. Kidney Int 1991; 39: 336–352.
Д.: Мочегонные средства. Kidney Int 1991; 39: 336–352. Совместная исследовательская группа DASH-Sodium. N Engl J Med 2001; 344: 3–10.
Совместная исследовательская группа DASH-Sodium. N Engl J Med 2001; 344: 3–10. Нат Жене 2003; 35: 372–376.
Нат Жене 2003; 35: 372–376. Б., Рикье А.Д., Пихакаски-Маунсбах К., Макдоноу А.А., Маунсбах А.Б.: Ангиотензин II провоцирует острую транспортировку котранспортера NaCl (NCC) из дистальных канальцев к апикальной мембране. Am J Physiol Renal Physiol 2007; 293: F662–F669.
Б., Рикье А.Д., Пихакаски-Маунсбах К., Макдоноу А.А., Маунсбах А.Б.: Ангиотензин II провоцирует острую транспортировку котранспортера NaCl (NCC) из дистальных канальцев к апикальной мембране. Am J Physiol Renal Physiol 2007; 293: F662–F669. Гипертония 2009;54:120–126.
Гипертония 2009;54:120–126. J Am Soc Nephrol 2011;22:460–471.
J Am Soc Nephrol 2011;22:460–471. Proc Natl Acad Sci USA 2006;103:1615–1620.
Proc Natl Acad Sci USA 2006;103:1615–1620. Hum Mol Genet 2011; 20: 855–866.
Hum Mol Genet 2011; 20: 855–866.
 гомеостаз.Proc Natl Acad Sci USA 2007;104:4025–4029.
гомеостаз.Proc Natl Acad Sci USA 2007;104:4025–4029. , Ривз В.Б., Рейли Р.Ф., Веласкес Х., Робертсон Н., Литвак Г., Морсинг П., Доруп Дж., Бахманн С., Эллисон Д.Х., Бостонджогло М.: экспрессия 11-бета-гидроксистероиддегидрогеназы, минералокортикоидного рецептора и тиазид-чувствительного котранспортера Na-Cl дистальными канальцами.J Am Soc Nephrol 1998; 9: 1347–1358.
, Ривз В.Б., Рейли Р.Ф., Веласкес Х., Робертсон Н., Литвак Г., Морсинг П., Доруп Дж., Бахманн С., Эллисон Д.Х., Бостонджогло М.: экспрессия 11-бета-гидроксистероиддегидрогеназы, минералокортикоидного рецептора и тиазид-чувствительного котранспортера Na-Cl дистальными канальцами.J Am Soc Nephrol 1998; 9: 1347–1358. , Ханссон Дж.Х., Шамбелан М., Гилл Дж.Р., Улик С., Милора Р.В., Финдлинг Дж.В., Канесса С.М., Россье Б.К., Лифтон Р.П.: Синдром Лиддла: наследственная человеческая гипертензия, вызванная мутациями в бета-субъединице эпителиального натриевого канала.Ячейка 1994; 79: 407–414.
, Ханссон Дж.Х., Шамбелан М., Гилл Дж.Р., Улик С., Милора Р.В., Финдлинг Дж.В., Канесса С.М., Россье Б.К., Лифтон Р.П.: Синдром Лиддла: наследственная человеческая гипертензия, вызванная мутациями в бета-субъединице эпителиального натриевого канала.Ячейка 1994; 79: 407–414.Автор Контакты
Gerardo Gamba
Vasco de Quiroga No. 15
Tlalpan, Mexico City 14000 (Мексика)
Тел. +52 55 5513 38 68
Электронная почта [email protected]
Информация о статье / публикации
Предварительный просмотр первой страницы
Получено: 19 января 2012 г.
Принято: 22 февраля 2012 г.
Опубликовано онлайн: 13 апреля 2012 г.
Дата выпуска выпуска: май 2012 г.
Количество печатных страниц: 8
Количество фигурок: 2
Количество столов: 1
ISSN: 0250-8095 (печать)
eISSN: 1421-9670 (онлайн)
Для получения дополнительной информации: https://www.karger.com/AJN
Авторское право / Дозировка препарата / Отказ от ответственности
Авторское право: Все права защищены. Никакая часть данной публикации не может быть переведена на другие языки, воспроизведена или использована в любой форме и любыми средствами, электронными или механическими, включая фотокопирование, запись, микрокопирование или любую систему хранения и поиска информации, без письменного разрешения издателя. .
Дозировка препарата: авторы и издатель приложили все усилия, чтобы гарантировать, что выбор препарата и дозировка, указанные в этом тексте, соответствуют текущим рекомендациям и практике на момент публикации. Тем не менее, в связи с продолжающимися исследованиями, изменениями в правительственных постановлениях и постоянным потоком информации, касающейся лекарственной терапии и реакций на лекарства, читателю настоятельно рекомендуется проверять вкладыш в упаковке для каждого лекарства на предмет любых изменений в показаниях и дозировке, а также для дополнительных предупреждений. и меры предосторожности. Это особенно важно, когда рекомендуемый агент является новым и/или редко используемым лекарственным средством.
Тем не менее, в связи с продолжающимися исследованиями, изменениями в правительственных постановлениях и постоянным потоком информации, касающейся лекарственной терапии и реакций на лекарства, читателю настоятельно рекомендуется проверять вкладыш в упаковке для каждого лекарства на предмет любых изменений в показаниях и дозировке, а также для дополнительных предупреждений. и меры предосторожности. Это особенно важно, когда рекомендуемый агент является новым и/или редко используемым лекарственным средством.
Отказ от ответственности: заявления, мнения и данные, содержащиеся в этой публикации, принадлежат исключительно отдельным авторам и участникам, а не издателям и редакторам.Появление рекламы и/или ссылок на продукты в публикации не является гарантией, одобрением или одобрением рекламируемых продуктов или услуг или их эффективности, качества или безопасности. Издатель и редактор(ы) отказываются от ответственности за любой ущерб людям или имуществу в результате любых идей, методов, инструкций или продуктов, упомянутых в содержании или рекламе.
: новая связь с гипертонией | Нефрология Диализная трансплантация
Путь почечной киназы WNK: новая связь с гипертонией
Открытие почечной киназы WNK позволяет по-новому взглянуть на регуляцию натрия, калия и артериального давления в дистальных отделах нефрона.Это также в значительной степени объяснило патогенез генетической формы гипертензии, называемой семейной гиперкалиемической гипертензией (СГГт, также известной как псевдогипоальдостеронизм II типа или синдром Гордона), поскольку она вызывается мутациями в киназах WNK. Однако возникает вопрос: имеют ли почечные киназы WNK клиническое значение помимо этого редкого синдрома? Здесь мы рассмотрим самые последние данные о физиологии почечных киназ WNK и обсудим их потенциально более широкую роль в транспорте электролитов и гипертонии.
Путь почечной киназы WNK: текущий статус
Как это часто бывает в науке, идентификация киназ WNK оказалась случайной находкой. В 2000 г. Сюй и др. использовали стратегию клонирования с вложенной полимеразной цепной реакцией для идентификации новых членов семейства митоген-активируемых белков/протеинкиназ, регулируемых внеклеточными сигналами [1]. Вместо этого они нашли нового члена семейства серин/треонинкиназ. Они назвали новую киназу WNK, что означает «без K», где «K» относится к аминокислоте лизину.Название подчеркивает их уникальную характеристику: расположение каталитического лизина, решающего для связывания с АТФ, в субдомене I, а не во II, как в случае всех других протеинкиназ. На сегодняшний день идентифицировано пять киназ WNK, а именно от WNK1 до WNK4 и специфичная для почек WNK1 (KS-WNK1, второй транскрипт гена WNK1). Физиологические функции киназ WNK разнообразны и включают регуляцию объема клеток, нейротрансмиссию, пролиферацию клеток, эмбриональное развитие, парацеллюлярную проницаемость и трансэпителиальный транспорт ионов [2].Сосредоточившись на последнем, было обнаружено, что киназы WNK занимают видное место во внутриклеточных сигнальных каскадах по всему нефрону, определяя активность или количество основных переносчиков натрия и калия в почках.
В 2000 г. Сюй и др. использовали стратегию клонирования с вложенной полимеразной цепной реакцией для идентификации новых членов семейства митоген-активируемых белков/протеинкиназ, регулируемых внеклеточными сигналами [1]. Вместо этого они нашли нового члена семейства серин/треонинкиназ. Они назвали новую киназу WNK, что означает «без K», где «K» относится к аминокислоте лизину.Название подчеркивает их уникальную характеристику: расположение каталитического лизина, решающего для связывания с АТФ, в субдомене I, а не во II, как в случае всех других протеинкиназ. На сегодняшний день идентифицировано пять киназ WNK, а именно от WNK1 до WNK4 и специфичная для почек WNK1 (KS-WNK1, второй транскрипт гена WNK1). Физиологические функции киназ WNK разнообразны и включают регуляцию объема клеток, нейротрансмиссию, пролиферацию клеток, эмбриональное развитие, парацеллюлярную проницаемость и трансэпителиальный транспорт ионов [2].Сосредоточившись на последнем, было обнаружено, что киназы WNK занимают видное место во внутриклеточных сигнальных каскадах по всему нефрону, определяя активность или количество основных переносчиков натрия и калия в почках. В дистальном отделе нефрона киназы WNK участвуют в регуляции Na-Cl-котранспортера в дистальных извитых канальцах (DCT) и эпителиальных натриевых каналах (ENaC) в соединительных канальцах (CNT) и собирательных трубочках (CD). Рисунок 1 ). В DCT содержание и фосфорилирование Na-Cl-котранспортера определяется соотношением между WNK3 (стимулятор) и WNK4 (ингибитор), т.е.е. они образуют «молекулярный реостат» [3]. Для дополнительной сложности WNK1 ингибирует WNK4, а KS-WNK1 ингибирует WNK1. Недавно также было показано, что STE20/SPS1-родственная пролин-аланин-богатая киназа и киназа-1, реагирующая на окислительный стресс (SPAK/OSR1), участвуют в регуляции Na-Cl-котранспортера [4] и, возможно, взаимодействуют с WNK3. и WNK4 [5]. В CNT и CD фосфорилирование ENaC регулируется WNK1 и KS-WNK1, но ингибируется WNK4. Киназа 1, регулируемая сывороткой и глюкокортикоидами (SGK1), играет центральную роль в этих эффектах, потому что WNK1 косвенно активирует ENaC за счет активации SGK1 через фосфатидилинозитол-3-киназу (PI3K), тогда как SGK1 обращает ингибирование ENaC с помощью WNK4.
В дистальном отделе нефрона киназы WNK участвуют в регуляции Na-Cl-котранспортера в дистальных извитых канальцах (DCT) и эпителиальных натриевых каналах (ENaC) в соединительных канальцах (CNT) и собирательных трубочках (CD). Рисунок 1 ). В DCT содержание и фосфорилирование Na-Cl-котранспортера определяется соотношением между WNK3 (стимулятор) и WNK4 (ингибитор), т.е.е. они образуют «молекулярный реостат» [3]. Для дополнительной сложности WNK1 ингибирует WNK4, а KS-WNK1 ингибирует WNK1. Недавно также было показано, что STE20/SPS1-родственная пролин-аланин-богатая киназа и киназа-1, реагирующая на окислительный стресс (SPAK/OSR1), участвуют в регуляции Na-Cl-котранспортера [4] и, возможно, взаимодействуют с WNK3. и WNK4 [5]. В CNT и CD фосфорилирование ENaC регулируется WNK1 и KS-WNK1, но ингибируется WNK4. Киназа 1, регулируемая сывороткой и глюкокортикоидами (SGK1), играет центральную роль в этих эффектах, потому что WNK1 косвенно активирует ENaC за счет активации SGK1 через фосфатидилинозитол-3-киназу (PI3K), тогда как SGK1 обращает ингибирование ENaC с помощью WNK4. Наконец, почечный наружный медуллярный калиевый канал (ROMK), который экспрессируется во всех этих сегментах нефрона, ингибируется WNK1 и WNK4, потому что они взаимодействуют с перекрестным каркасным белком, чтобы стимулировать клатрин-опосредованный эндоцитоз [6]. За последние 8 лет исследования с использованием ооцитов Xenopus и клеток HEK-293 сыграли важную роль в раскрытии молекулярного механизма пути почечной киназы WNK. В настоящее время первые исследования на животных сосредоточены на взаимосвязи между альдостероном и киназами WNK.Было показано, что у мышей и крыс дикого типа нагрузка калием и альдостероном увеличивает мРНК KS-WNK1 и WNK4 [7,8], тогда как ограничение калия снижает KS-WNK1 и увеличивает WNK1 [9]. Хотя мРНК KS-WNK1 была значительно выше у мышей на диете с высоким содержанием натрия по сравнению с мышами на диете с низким содержанием натрия [7], никакой разницы в уровне белка KS-WNK1 или WNK4 не наблюдалось [4]. Эти результаты могут пролить свет на так называемый «парадокс альдостерона», то есть на давний физиологический вопрос о том, как альдостерон может быть гормоном, удерживающим натрий, и гормоном, секретирующим калий [2, 10].
Наконец, почечный наружный медуллярный калиевый канал (ROMK), который экспрессируется во всех этих сегментах нефрона, ингибируется WNK1 и WNK4, потому что они взаимодействуют с перекрестным каркасным белком, чтобы стимулировать клатрин-опосредованный эндоцитоз [6]. За последние 8 лет исследования с использованием ооцитов Xenopus и клеток HEK-293 сыграли важную роль в раскрытии молекулярного механизма пути почечной киназы WNK. В настоящее время первые исследования на животных сосредоточены на взаимосвязи между альдостероном и киназами WNK.Было показано, что у мышей и крыс дикого типа нагрузка калием и альдостероном увеличивает мРНК KS-WNK1 и WNK4 [7,8], тогда как ограничение калия снижает KS-WNK1 и увеличивает WNK1 [9]. Хотя мРНК KS-WNK1 была значительно выше у мышей на диете с высоким содержанием натрия по сравнению с мышами на диете с низким содержанием натрия [7], никакой разницы в уровне белка KS-WNK1 или WNK4 не наблюдалось [4]. Эти результаты могут пролить свет на так называемый «парадокс альдостерона», то есть на давний физиологический вопрос о том, как альдостерон может быть гормоном, удерживающим натрий, и гормоном, секретирующим калий [2, 10]. А именно, если нагрузка калием (гиперкалиемия) активирует KS-WNK1 и WNK4, это будет ингибировать котранспортер Na-Cl и способствовать электрогенной реабсорбции натрия с помощью ENaC, тем самым увеличивая трансэпителиальный потенциал и стимулируя секрецию калия. Обратное происходит, когда диета с низким содержанием натрия (гиповолемия) не влияет или даже снижает KS-WNK1 и WNK4, потому что это активирует Na-Cl-котранспортер и способствует электронейтральной реабсорбции натрия при относительном сохранении калия.
А именно, если нагрузка калием (гиперкалиемия) активирует KS-WNK1 и WNK4, это будет ингибировать котранспортер Na-Cl и способствовать электрогенной реабсорбции натрия с помощью ENaC, тем самым увеличивая трансэпителиальный потенциал и стимулируя секрецию калия. Обратное происходит, когда диета с низким содержанием натрия (гиповолемия) не влияет или даже снижает KS-WNK1 и WNK4, потому что это активирует Na-Cl-котранспортер и способствует электронейтральной реабсорбции натрия при относительном сохранении калия.
Рис. 1
Путь почечной киназы WNK в дистальном отделе нефрона. Текущая модель пути почечной киназы WNK в дистальном отделе нефрона, включая эффекты альдостерона, диеты с низким содержанием калия (низкий K + ) и инсулина. Стимулирующие эффекты показаны красными стрелками с символом «+», тормозные эффекты показаны черными стрелками с символом «-». Фосфорилирование обозначается символом «P».
Рис. 1
Путь почечной киназы WNK в дистальном отделе нефрона. Текущая модель пути почечной киназы WNK в дистальном отделе нефрона, включая эффекты альдостерона, диеты с низким содержанием калия (низкий K + ) и инсулина. Стимулирующие эффекты показаны красными стрелками с символом «+», тормозные эффекты показаны черными стрелками с символом «-». Фосфорилирование обозначается символом «P».
Текущая модель пути почечной киназы WNK в дистальном отделе нефрона, включая эффекты альдостерона, диеты с низким содержанием калия (низкий K + ) и инсулина. Стимулирующие эффекты показаны красными стрелками с символом «+», тормозные эффекты показаны черными стрелками с символом «-». Фосфорилирование обозначается символом «P».
WNK-киназы и гипертензия
Киназы WNK привлекли наибольшее внимание как причину FHHt.Исследования позиционного клонирования пациентов с FHHt выявили две причины [11]: интронные делеции, вызывающие сверхэкспрессию WNK1 дикого типа (которая ингибирует WNK4), и миссенс-мутации, вызывающие мутантную WNK4 (которая ингибирует WNK4 дикого типа и не ингибирует WNK3). Мыши, трансгенные по мутантному WNK4 [12], и нокаутированные мыши с одним мутантным и одним аллелем WNK4 дикого типа [13] повторяют фенотип FHHt. Редкость FHHt поднимает вопрос, имеют ли киназы WNK клиническую значимость помимо этого синдрома. Чтобы ответить на этот вопрос, мы сначала рассмотрим два других вопроса: является ли нынешнее лечение артериальной гипертензии неоптимальным, и если да, то что необходимо для его улучшения? По нашему мнению, ответ на первый вопрос однозначно «да», потому что около 90% случаев артериальной гипертензии по-прежнему считаются «необходимыми», а лечение в основном проводится методом проб и ошибок. Ответ на второй вопрос логично вытекает из первого: для лучшего ведения АГ необходимо лучшее понимание ее патогенеза и маркеров для определения индивидуальной чувствительности больного к антигипертензивным препаратам.Могут ли почечные киназы WNK способствовать этой миссии? Мы так считаем и приводим три примера. Во-первых, почечный путь киназы WNK предлагает потенциальное механистическое объяснение связи между истощением калия и солечувствительной гипертензией [14]. Наш рацион постепенно изменился от богатого калием и бедного натрием во времена палеолита до противоположного в наше время [15]. Как уже отмечалось, ограничение калия увеличивает WNK1 и снижает KS-WNK1 у животных [7-9].
Чтобы ответить на этот вопрос, мы сначала рассмотрим два других вопроса: является ли нынешнее лечение артериальной гипертензии неоптимальным, и если да, то что необходимо для его улучшения? По нашему мнению, ответ на первый вопрос однозначно «да», потому что около 90% случаев артериальной гипертензии по-прежнему считаются «необходимыми», а лечение в основном проводится методом проб и ошибок. Ответ на второй вопрос логично вытекает из первого: для лучшего ведения АГ необходимо лучшее понимание ее патогенеза и маркеров для определения индивидуальной чувствительности больного к антигипертензивным препаратам.Могут ли почечные киназы WNK способствовать этой миссии? Мы так считаем и приводим три примера. Во-первых, почечный путь киназы WNK предлагает потенциальное механистическое объяснение связи между истощением калия и солечувствительной гипертензией [14]. Наш рацион постепенно изменился от богатого калием и бедного натрием во времена палеолита до противоположного в наше время [15]. Как уже отмечалось, ограничение калия увеличивает WNK1 и снижает KS-WNK1 у животных [7-9]. Снижение KS-WNK1 ослабит его ингибирование WNK1, позволяя ему либо ингибировать WNK4 и активировать Na-Cl-котранспортер (DCT), либо активировать SGK1 и ENaC (CNT и CD, рисунок 1).В результате увеличивается реабсорбция натрия в двух участках нефрона, что приводит к повышению артериального давления. Вторым примером является связь между гипертензией и гиперинсулинемией, характерная черта таких заболеваний, как сахарный диабет и ожирение. Песня и др. показали, что повышение артериального давления у крыс при длительном лечении инсулином, вероятно, было связано с усилением реабсорбции натрия Na-Cl-котранспортером и ENaC, поскольку апикальная локализация субъединиц ENaC увеличивалась, а лечение гидрохлоротиазидом и амилоридом приводило к усилению натрийуреза. 16].Интересно, что инсулин снижал корковую экспрессию WNK4 [16], что действительно должно было активировать Na-Cl-котранспортер и ENaC (рис. 1). Третий пример заключается в том, что одиночные нуклеотидные полиморфизмы и гаплотипы в WNK1 вносят вклад в изменение артериального давления в общей популяции [17], возможно, опосредованное влиянием на градиент изменения артериального давления с возрастом [18].
Снижение KS-WNK1 ослабит его ингибирование WNK1, позволяя ему либо ингибировать WNK4 и активировать Na-Cl-котранспортер (DCT), либо активировать SGK1 и ENaC (CNT и CD, рисунок 1).В результате увеличивается реабсорбция натрия в двух участках нефрона, что приводит к повышению артериального давления. Вторым примером является связь между гипертензией и гиперинсулинемией, характерная черта таких заболеваний, как сахарный диабет и ожирение. Песня и др. показали, что повышение артериального давления у крыс при длительном лечении инсулином, вероятно, было связано с усилением реабсорбции натрия Na-Cl-котранспортером и ENaC, поскольку апикальная локализация субъединиц ENaC увеличивалась, а лечение гидрохлоротиазидом и амилоридом приводило к усилению натрийуреза. 16].Интересно, что инсулин снижал корковую экспрессию WNK4 [16], что действительно должно было активировать Na-Cl-котранспортер и ENaC (рис. 1). Третий пример заключается в том, что одиночные нуклеотидные полиморфизмы и гаплотипы в WNK1 вносят вклад в изменение артериального давления в общей популяции [17], возможно, опосредованное влиянием на градиент изменения артериального давления с возрастом [18]. Интересно, что ген, кодирующий SPAK ( STK39 ), который взаимодействует с киназами WNK (рис. 1), также недавно был идентифицирован как ген предрасположенности к гипертонии в популяции амишей [19].Возможность предсказывать индивидуальную предрасположенность к АГ логично приводит к последнему оставшемуся без ответа вопросу: существуют ли биомаркеры АГ? Скрининг полиморфизмов и гаплотипов может стать полезным клиническим инструментом в обозримом будущем, хотя доступные тесты не нашли широкого применения в клинической практике [20]. Было трудно установить четкие связи между полиморфизмами артериального давления (например, α-аддуцин, ангиотензиноген, ангиотензинпревращающий фермент) и риском гипертонии, сердечно-сосудистых событий или реакции на терапию [21,22].Однако скромный эффект вариантов в одном гене можно объяснить их взаимодействием с родственными генами. Действительно, при комбинировании вариантов в генах WNK1, α-аддуцина (влияет на активность Na-K-АТФазы) и Nedd4-2 (убихинаты ENaC) был обнаружен значительный эффект на почечную обработку соли, реакцию артериального давления на физиологический раствор и тиазиды и ночное систолическое артериальное давление [23].
Интересно, что ген, кодирующий SPAK ( STK39 ), который взаимодействует с киназами WNK (рис. 1), также недавно был идентифицирован как ген предрасположенности к гипертонии в популяции амишей [19].Возможность предсказывать индивидуальную предрасположенность к АГ логично приводит к последнему оставшемуся без ответа вопросу: существуют ли биомаркеры АГ? Скрининг полиморфизмов и гаплотипов может стать полезным клиническим инструментом в обозримом будущем, хотя доступные тесты не нашли широкого применения в клинической практике [20]. Было трудно установить четкие связи между полиморфизмами артериального давления (например, α-аддуцин, ангиотензиноген, ангиотензинпревращающий фермент) и риском гипертонии, сердечно-сосудистых событий или реакции на терапию [21,22].Однако скромный эффект вариантов в одном гене можно объяснить их взаимодействием с родственными генами. Действительно, при комбинировании вариантов в генах WNK1, α-аддуцина (влияет на активность Na-K-АТФазы) и Nedd4-2 (убихинаты ENaC) был обнаружен значительный эффект на почечную обработку соли, реакцию артериального давления на физиологический раствор и тиазиды и ночное систолическое артериальное давление [23]. При этом гены все еще далеки от реальной биологической рабочей силы, а именно белков. Следовательно, хотелось бы измерить активность Na-Cl-котранспортера, ENaC или WNK в дистальном отделе нефрона.Поскольку биопсия почки не проводится регулярно у пациентов с артериальной гипертензией, логичной альтернативой может быть моча, так как она содержит много белков, ассоциированных с заболеванием. Исследования, подтверждающие этот принцип, продемонстрировали повышенную экскрецию с мочой Na-Cl-котранспортера у пациентов с СГГТ [24] и специфический характер мочеиспускания простазина-активатора ENaC у пациентов с первичным альдостеронизмом [25]. В этих исследованиях использовалась цельная моча, но более целенаправленным подходом могло бы быть использование так называемых мочевых экзосом.Мочевые экзосомы представляют собой внутренние везикулы мультивезикулярных телец, секретируемые почечными эпителиальными клетками и содержащие Na-Cl-котранспортер и ENaC (неизвестно, присутствуют ли в экзосомах киназы WNK) [26].
При этом гены все еще далеки от реальной биологической рабочей силы, а именно белков. Следовательно, хотелось бы измерить активность Na-Cl-котранспортера, ENaC или WNK в дистальном отделе нефрона.Поскольку биопсия почки не проводится регулярно у пациентов с артериальной гипертензией, логичной альтернативой может быть моча, так как она содержит много белков, ассоциированных с заболеванием. Исследования, подтверждающие этот принцип, продемонстрировали повышенную экскрецию с мочой Na-Cl-котранспортера у пациентов с СГГТ [24] и специфический характер мочеиспускания простазина-активатора ENaC у пациентов с первичным альдостеронизмом [25]. В этих исследованиях использовалась цельная моча, но более целенаправленным подходом могло бы быть использование так называемых мочевых экзосом.Мочевые экзосомы представляют собой внутренние везикулы мультивезикулярных телец, секретируемые почечными эпителиальными клетками и содержащие Na-Cl-котранспортер и ENaC (неизвестно, присутствуют ли в экзосомах киназы WNK) [26]. Мочевые экзосомы не анализировались при гипертензивных расстройствах, но их полезность иллюстрируется идентификацией экзосомальных биомаркеров, которые способны предсказывать острую почечную недостаточность до повышения уровня креатинина в сыворотке [27,28].
Мочевые экзосомы не анализировались при гипертензивных расстройствах, но их полезность иллюстрируется идентификацией экзосомальных биомаркеров, которые способны предсказывать острую почечную недостаточность до повышения уровня креатинина в сыворотке [27,28].
Перспективы
Роль почечных киназ WNK и их взаимодействия с переносчиками натрия и калия в быстро развивающихся клеточных моделях DCT, CNT и CD становится все более ясной.Тем не менее роль WNK3 и особенно WNK2 в дистальных отделах нефрона относительно неизвестна. Кроме того, биология никогда не бывает такой простой, как одно семейство белков, и, по-видимому, по крайней мере три киназные системы координируют передачу сигнала от рецептора к транспортеру, включая киназы WNK, SGK1 и SPAK/OSR1 (Рис. 1). Хотя альдостерон является бесспорным активатором киназ WNK, неизвестно, способны ли другие гормоны, действующие на дистальные отделы нефрона, такие как вазопрессин, ангиотензин II и предсердный натрийуретический пептид, также регулировать киназы WNK. Первые исследования на животных были сосредоточены на альдостероне и киназах WNK, но полная картина, вероятно, также требует анализа других циркулирующих гормонов, связанных рецепторов и транспортеров и, конечно же, артериального давления. Помимо физиологических данных, кажется логичным продолжить поиск мочевых биомаркеров гипертонии [29]. На данный момент использование почечных WNK-киназ в качестве мишеней для лекарств является научной фантастикой, но пример ингибитора тирозинкиназы иматиниба для лечения хронического миелоидного лейкоза показывает, что селективное ингибирование киназных систем не является априорно невозможным [30,31].Возможность ингибирования WNK1 также была продемонстрирована на мышах, гетерозиготных по мутации WNK1, у которых наблюдалось заметное снижение артериального давления без явных побочных эффектов [32]. Гипертония, очевидно, является многофакторным и сложным заболеванием, но путь киназы WNK открывает привлекательный путь для лучшего понимания и потенциальной диагностики и лечения гипертонии.
Первые исследования на животных были сосредоточены на альдостероне и киназах WNK, но полная картина, вероятно, также требует анализа других циркулирующих гормонов, связанных рецепторов и транспортеров и, конечно же, артериального давления. Помимо физиологических данных, кажется логичным продолжить поиск мочевых биомаркеров гипертонии [29]. На данный момент использование почечных WNK-киназ в качестве мишеней для лекарств является научной фантастикой, но пример ингибитора тирозинкиназы иматиниба для лечения хронического миелоидного лейкоза показывает, что селективное ингибирование киназных систем не является априорно невозможным [30,31].Возможность ингибирования WNK1 также была продемонстрирована на мышах, гетерозиготных по мутации WNK1, у которых наблюдалось заметное снижение артериального давления без явных побочных эффектов [32]. Гипертония, очевидно, является многофакторным и сложным заболеванием, но путь киназы WNK открывает привлекательный путь для лучшего понимания и потенциальной диагностики и лечения гипертонии.
EJH поддерживается стипендией Erasmus MC Fellowship 2008 (внутренний грант) и грантом Kolff Junior Postdoc (Голландский почечный фонд).
Заявление о конфликте интересов . Ни один не заявил.
Ссылки
1, , , и др.WNK1, новая серин/треониновая протеинкиназа млекопитающих, в которой отсутствует каталитический лизин в субдомене II
275
стр.16795
2, , .Киназы WNK и почечный транспорт натрия в норме и при патологии: комплексный взгляд
51
стр.588
3, , .Чувствительный к тиазидам котранспортер Na-Cl регулируется сигнальным комплексом киназы WNK
,J Clin Invest
,2007
, vol.117
стр.3403
4, , , и др.Пищевая соль регулирует фосфорилирование киназ OSR1/SPAK и котранспортера хлорида натрия через альдостерон
74
стр.1403
5.Регуляция котранспортера Na + -Cl − с помощью пищевого NaCl: роль WNK, SPAK, OSR1 и альдостерона
74
стр.
1373
6, , .Механизм регуляции почечного транспорта ионов киназами WNK
17
стр.519
7, , , и др.Пищевые электролитные ответы в пути почечной киназы WNK in vivo
,J Am Soc Nephrol
,2006
, vol.17
стр.2402
8, , , и др.Переключатель изоформы киназы WNK1 регулирует экскрецию калия почками
103
стр.8558
9, , .Антагонистическая регуляция ROMK длинными и специфичными для почек изоформами WNK1
103
стр.1615
10, , .Синдромы Барттера, Гительмана и Гордона. От физиологии к молекулярной биологии и обратно, но еще вопросы без ответа
,Нефрон
,2002
, том.92
Дополнение 1
стр.18
11, , , и др.Гипертензия человека, вызванная мутациями киназ WNK
,Science
,2001
, vol.293
стр.1107
12, , , и др.Wnk4 контролирует кровяное давление и гомеостаз калия посредством регуляции массы и активности дистального извитого канальца
38
стр.
1124
13, , , и др.Молекулярный патогенез псевдогипоальдостеронизма типа II: создание и анализ мышиной модели с нокаутом Wnk4(D561A/+)
5
стр.331
14, .Механизмы заболевания: WNK-инг в механизме солевой гипертензии
,Nat Clin Pract Nephrol
,2007
, том.3
стр.623
15, , , и др.Контроль экскреции калия: палеолитическая перспектива
15
стр.430
16, , , и др.Регуляция артериального давления, эпителиального натриевого канала (ENaC) и других ключевых переносчиков натрия в почках путем хронической инфузии инсулина у крыс
290
стр.F1055
17, , , и др.Связь полиморфизмов и гаплотипов гена WNK1 с амбулаторным артериальным давлением в общей популяции
,Тираж
,2005
, том112
стр.3423
18, , , и др.Общие вариации гена WNK1 и артериального давления в детском возрасте: Продольное исследование родителей и детей Avon
52
стр.
974
19, , , и др.Полногеномное ассоциативное исследование идентифицирует STK39 как ген предрасположенности к артериальной гипертензии
43
стр.1155
21, , , и др.Полиморфизм I/D гена ACE не связан с преимуществами ингибирования ACE для артериального давления и сердечно-сосудистой системы
42
стр.297
22, , , и др.Однонуклеотидные полиморфизмы ангиотензиногена, повышенное кровяное давление и риск сердечно-сосудистых заболеваний
41
стр.1202
23, , , и др.Физиологическое взаимодействие между путями альфа-аддуцина и WNK1-NEDD4L в регуляции артериального давления, связанной с натрием
52
стр.366
24, , , и др.Повышенное содержание белка-котранспортера Na-Cl в моче при семейной гиперкалиемии и гипертензии
23
стр.492
25, , , и др.Мочевой простазин: кандидатный маркер активации эпителиальных натриевых каналов у человека
46
стр.683
26, , .
Идентификация и протеомное профилирование экзосом в моче человека
101
стр.13368
27, , , и др.Факторы экзосомальной транскрипции мочи, новый класс биомаркеров почечной недостаточности
,Kidney Int
,2008
, том.74
стр.613
28, , , и др.Экзосомальный фетуин-А, идентифицированный с помощью протеомики: новый биомаркер мочи для выявления острого повреждения почек
70
стр.1847
29, , , и др.Перспективы протеомики мочи: экзосомы как источник биомаркеров мочи
10
стр.283
30, , .Киназы WNK и контроль артериального давления
,Pharmacol Ther
,2005
, vol.106
стр.221
31, , .Ингибиторы протеинкиназы: взгляд на дизайн лекарств из структуры
,Наука
,2004
, том.303
стр.1800
32, , , и др.Дефицит киназы Wnk1 снижает артериальное давление у мышей: скрининг генных ловушек для выявления потенциальных мишеней для терапевтического вмешательства
100
стр.
14109
© Автор [2009]. Опубликовано Oxford University Press от имени ERA-EDTA. Все права защищены. Для получения разрешений обращайтесь по электронной почте: [email protected]
Oxford University Press
Сигнальные пути WNK в регуляции кровяного давления
Нараян К.М., Али М.К., Коплан Дж.П. (2010) Глобальные неинфекционные заболевания — место встречи миров. N Engl J Med 363 (13): 1196–1198.дои: 10.1056/NEJMp1002024
КАС пабмед Статья Google Scholar
Guyton AC (1991) Контроль артериального давления – особая роль почек и жидкостей организма. Science 252 (5014): 1813–1816
CAS пабмед Статья Google Scholar
Paver WK, Pauline GJ (1964) Гипертензия и гиперкалиемия без почечной недостаточности у молодого мужчины. Med J Aust 2:305–306
Med J Aust 2:305–306
CAS пабмед Google Scholar
Mayan H, Vered I, Mouallem M, Tzadok-Witkon M, Pauzner R, Farfel Z (2002) Псевдогипоальдостеронизм II типа: выраженная чувствительность к тиазидам, гиперкальциурия, нормомагниемия и низкая минеральная плотность костей. J Clin Endocrinol Metab 87(7):3248–3254. doi:10.1210/jcem.87.7.8449
КАС пабмед Статья Google Scholar
Simon DB, Nelson-Williams C, Bia MJ, Ellison D, Karet FE, Molina AM, Vaara I, Iwata F, Cushner HM, Koolen M, Gainza FJ, Gitleman HJ, Lifton RP (1996) Гительмановский вариант синдрома Барттера, наследственный гипокалиемический алкалоз вызывается мутациями в тиазид-чувствительном котранспортере Na-Cl. Нат Жене 12 (1): 24–30. дои: 10.1038/ng0196-24
КАС пабмед Статья Google Scholar

Мэннинг Г., Уайт Д.Б., Мартинес Р., Хантер Т., Сударсанам С. (2002) Протеинкиназный комплемент человеческого генома.Наука 298 (5600): 1912–1934. дои: 10.1126/наука.1075762
КАС пабмед Статья Google Scholar
Piala AT, Moon TM, Akella R, He H, Cobb MH, Goldsmith EJ (2014) Восприятие хлорида киназой WNK1 включает ингибирование аутофосфорилирования. Научный сигнал 7 (324): ra41. doi: 10.1126/scisignal.2005050
Lenertz LY, Lee BH, Min X, B-e Xu, Wedin K, Earnest S, Goldsmith EJ, Cobb MH (2005) Свойства WNK1 и последствия для других членов семьи.J Biol Chem 280(29):26653–26658. дои: 10.1074/jbc.M502598200
КАС пабмед Статья Google Scholar
Richardson C, Alessi DR (2008) Регуляция транспорта соли и артериального давления сигнальным путем WNK-SPAK/OSR1. J Cell Sci 121 (Pt 20): 3293–3304. дои: 10.1242/jcs.029223
J Cell Sci 121 (Pt 20): 3293–3304. дои: 10.1242/jcs.029223
КАС пабмед Статья Google Scholar
Zagórska A, Pozo-Guisado E, Boudeau J, Vitari AC, Rafiqi FH, Thastrup J, Deak M, Campbell DG, Morrice NA, Prescott AR, Alessi DR (2007) Регуляция активности и локализации протеинкиназы WNK1 с помощью гиперосмотической стресс. J Cell Biol 176 (1): 89–100. дои: 10.1083/jcb.200605093
ПабМед ПабМед Центральный Статья КАС Google Scholar
Haas M, Forbush B 3rd (2000) Котранспортер Na-K-Cl секреторного эпителия.Annu Rev Physiol 62: 515–534. doi:10.1146/annurev.physiol.62.1.515
КАС пабмед Статья Google Scholar
Kahle KT, Rinehart J, Lifton RP (2010) Фосфорегуляция котранспортеров Na-K-2Cl и K-Cl с помощью киназ WNK. Биохим Биофиз Акта 1802 (12): 1150–1158. doi:10.1016/j.bbadis.2010.07.009
Биохим Биофиз Акта 1802 (12): 1150–1158. doi:10.1016/j.bbadis.2010.07.009
КАС пабмед ПабМед Центральный Статья Google Scholar
B-e Xu, English JM, Wilsbacher JL, Stippec S, Goldsmith EJ, Cobb MH (2000) WNK1, новая серин/треониновая протеинкиназа млекопитающих, в которой отсутствует каталитический лизин в субдомене II. J Biol Chem 275(22):16795–16801. дои: 10.1074/jbc.275.22.16795
Артикул Google Scholar
Уилсон Ф.Х., Диссе-Никодем С., Чоат К.А., Исикава К., Нельсон-Уильямс С., Дезиттер И., Гюнель М., Милфорд Д.В., Липкин Г.В., Ачард Дж.М., Фили М.П., Дюссол Б., Берланд И., Анвин RJ, Mayan H, Simon DB, Farfel Z, Jeunemaitre X, Lifton RP (2001)Гипертензия человека, вызванная мутациями в киназах WNK.Наука 293 (5532): 1107–1112. дои: 10.1126/наука.1062844
КАС пабмед Статья Google Scholar

Бойден Л.М., Чой М., Чоат К.А., Нельсон-Уильямс С.Дж., Фархи А., Тока Х.Р., Тихонова И.Р., Бьорнсон Р., Мане С.М., Колусси Г., Лебель М., Гордон Р.Д., Семмикрот Б.А., Пужол А., Валимаки М.Дж., Де Феррари М.Е., Санджад С.А., Гуткин М., Карет Ф.Е., Туччи М.Р., Стокигт М.Р., Кепплер-Норей К.М., Портер С.К., Ананд С.К., Уайтфорд М.Л., Дэвис И.Д., Дьюар С.Б., Беттинелли А., Фадровски Д.Дж., Белша К.В. , Ханли Т.Е., Нельсон Р.Д., Трахтман Х., Коул Т.Р., Пинск М., Бокенхауэр Д., Шеной М., Вайдьянатан П., Форман Дж.В., Расулпур М., Тамим Ф., Аль-Шахрури Х.З., Радхакришнан Дж., Гарави А.Г., Гойлав Б., Лифтон Р.П. (2012) Мутации в kelch-like 3 и cullin 3 вызывают гипертензию и нарушения электролитного баланса.Природа 482 (7383): 98–102. дои: 10.1038 / природа10814
КАС пабмед ПабМед Центральный Статья Google Scholar
Шумахер Ф.Р., Сью К., Чжан Дж., Джонсон С., Вуд Н., Клири С.Э., Аль Маскари Р. С., Ферриман Дж.Т., Хардедж И., Ясмин Фигг Н.Л., Энчев Р., Кнебель А., О’Шонесси К.М., Курц Т (2015) Характеристика мутации куллина-3, которая вызывает тяжелую форму семейной гипертензии и гиперкалиемии.EMBO Mol Med 7 (10): 1285–1306. дои: 10.15252/эммм.201505444
С., Ферриман Дж.Т., Хардедж И., Ясмин Фигг Н.Л., Энчев Р., Кнебель А., О’Шонесси К.М., Курц Т (2015) Характеристика мутации куллина-3, которая вызывает тяжелую форму семейной гипертензии и гиперкалиемии.EMBO Mol Med 7 (10): 1285–1306. дои: 10.15252/эммм.201505444
КАС пабмед ПабМед Центральный Статья Google Scholar
Ohta A, Schumacher FR, Mehellou Y, Johnson C, Knebel A, Macartney TJ, Wood NT, Alessi DR, Kurz T (2013) Комплекс лигазы cul3–klhl3 e3, мутировавший при синдроме гипертензии Гордона, взаимодействует с и убиквитилирует изоформы WNK: вызывающие заболевание мутации в KLHL3 и WNK4 нарушают взаимодействие.Биохим J 451 (1): 111–122. дои: 10.1042/bj20121903
КАС пабмед ПабМед Центральный Статья Google Scholar
Луи-Дит-Пикард Х., Барк Дж., Трухильяно Д., Мизерей-Ленкей С. , Буатиа-Наджи Н., Пилипенко О., Борен Г., Боннефон А., Санд О., Симиан С., Видаль-Петио Э., Сукасеум С., Манде С., Брукс Ф., Шабр О., Делаусс М., Эсно В., Фике Б., Уилье П., Баньис С.И., Кениг Дж., Конрад М., Ландаис П., Мурани С., Ниодет П., Пробст В., Товен С., Унвин Р.Дж., Soroka SD, Ehret G, Ossowski S, Caulfield M, International Consortium for Blood P, Bruneval P, Estivill X, Froguel P, Hadchouel J, Schott JJ, Jeunemaitre X (2012) Мутации KLHL3 вызывают семейную гиперкалиемическую гипертензию, нарушая транспорт ионов в дистальный отдел нефрона.Nat Genet 44 (4): 456–460 (S451–S453). дои: 10.1038 / ng.2218
, Буатиа-Наджи Н., Пилипенко О., Борен Г., Боннефон А., Санд О., Симиан С., Видаль-Петио Э., Сукасеум С., Манде С., Брукс Ф., Шабр О., Делаусс М., Эсно В., Фике Б., Уилье П., Баньис С.И., Кениг Дж., Конрад М., Ландаис П., Мурани С., Ниодет П., Пробст В., Товен С., Унвин Р.Дж., Soroka SD, Ehret G, Ossowski S, Caulfield M, International Consortium for Blood P, Bruneval P, Estivill X, Froguel P, Hadchouel J, Schott JJ, Jeunemaitre X (2012) Мутации KLHL3 вызывают семейную гиперкалиемическую гипертензию, нарушая транспорт ионов в дистальный отдел нефрона.Nat Genet 44 (4): 456–460 (S451–S453). дои: 10.1038 / ng.2218
Kahle KT, Wilson FH, Leng Q, Lalioti MD, O’Connell AD, Dong K, Rapson AK, MacGregor GG, Giebisch G, Hebert SC, Lifton RP (2003) WNK4 регулирует баланс между почечным NaCl реабсорбции и секреции К+. Нат Жене 35 (4): 372–376. дои: 10.1038/ng1271
КАС пабмед Статья Google Scholar

Kahle KT, Macgregor GG, Wilson FH, Van Hoek AN, Brown D, Ardito T, Kashgarian M, Giebisch G, Hebert SC, Boulpaep EL, Lifton RP (2004) Парацеллюлярная Cl-проницаемость регулируется киназой WNK4: понимание нормального физиология и артериальная гипертензия. Proc Natl Acad Sci USA 101(41):14877–14882. doi:10.1073/pnas.0406172101
КАС пабмед ПабМед Центральный Статья Google Scholar
Melo Z, Cruz-Rangel S, Bautista R, Vazquez N, Castaneda-Bueno M, Mount DB, Pasantes-Morales H, Mercado A, Gamba G (2013) Молекулярные доказательства роли K(+ )-Cl(-) котранспортеры в почках.Am J Physiol Renal Physiol 305(10):F1402–F1411. doi:10.1152/ajprenal.00390.2013
КАС пабмед ПабМед Центральный Статья Google Scholar
Yang CL, Angell J, Mitchell R, Ellison DH (2003) Киназы WNK регулируют тиазид-чувствительный котранспорт Na-Cl. J Clin Invest 111(7):1039–1045
J Clin Invest 111(7):1039–1045
CAS пабмед ПабМед Центральный Статья Google Scholar
Leng Q, Kahle KT, Rinehart J, MacGregor GG, Wilson FH, Canessa CM, Lifton RP, Hebert SC (2006) WNK3, киназа, родственная генам, мутировавшим при наследственной гипертензии с гиперкалиемией, регулирует K+ канал ROMK1 (kir1.1 ). J Physiol 571 (Pt 2): 275–286. doi:10.1113/jphysiol.2005.102202
Naray-Fejes-Toth A, Snyder PM, Fejes-Toth G (2004) Специфическая для почек изоформа WNK1 индуцируется альдостероном и стимулирует перенос Na+ через эпителиальные натриевые каналы.Proc Natl Acad Sci USA 101(50):17434–17439. doi:10.1073/pnas.0408146101
КАС
пабмед
ПабМед Центральный
Статья
Google Scholar
(2013) ) Семейная гиперкалиемическая гипертензия, связанная с WNK1, возникает в результате повышенной экспрессии l-WNK1, особенно в дистальных отделах нефрона. Proc Natl Acad Sci USA 110(35):14366–14371. doi:10.1073/pnas.1304230110
Proc Natl Acad Sci USA 110(35):14366–14371. doi:10.1073/pnas.1304230110
КАС пабмед ПабМед Центральный Статья Google Scholar
Лифтон Р.П., Гарави А.Г., Геллер Д.С. (2001) Молекулярные механизмы гипертонии человека. Сотовый 104(4):545–556
CAS пабмед Статья Google Scholar
Na T, Wu G, Zhang W, Dong WJ, Peng JB (2013) Болезнетворная мутация r1185c WNK4 нарушает регуляторный механизм, включающий сайты связывания кальмодулина и фосфорилирования SGK1.Am J Physiol Renal Physiol 304(1):F8–F18. doi: 10.1152/ajprenal.00284.2012
КАС пабмед Статья Google Scholar
Arroyo JP, Kahle KT, Gamba G (2013) Семейство slc12 электронейтральных переносчиков хлоридов, связанных с катионами. Mol Aspects Med 34 (2–3): 288–298. doi: 10.1016 / j.mam.2012.05.002
doi: 10.1016 / j.mam.2012.05.002
КАС пабмед Статья Google Scholar
Gamba G (2005)Молекулярная физиология и патофизиология электронейтральных катион-хлоридных котранспортеров. Physiol Rev 85 (2): 423–493. doi:10.1152/physrev.00011.2004
КАС пабмед Статья Google Scholar
Strange K, Denton J, Nehrke K (2006)Киназы типа Ste20: эволюционно консервативные регуляторы транспорта ионов и объема клеток. Физиология (Ветесда) 21:61–68. doi:10.1152/физиол.00139.2005
КАС Статья Google Scholar
Delaloy C, Lu J, Houot AM, Disse-Nicodeme S, Gasc JM, Corvol P, Jeunemaitre X (2003) Множественные промоторы в гене WNK1: один контролирует экспрессию специфичной для почек изоформы, дефектной по киназы . Mol Cell Biol 23(24):9208–9221
Mol Cell Biol 23(24):9208–9221
CAS пабмед ПабМед Центральный Статья Google Scholar
O’Reilly M, Marshall E, Speirs HJ, Brown RW (2003) WNK1, ген нового пути контроля артериального давления, тканеспецифически генерирует радикально отличающиеся изоформы с киназным доменом и без него. J Am Soc Nephrol 14(10):2447–2456
PubMed Статья КАС Google Scholar
Wilson FH, Kahle KT, Sabath E, Lalioti MD, Rapson AK, Hoover RS, Hebert SC, Gamba G, Lifton RP (2003) Молекулярный патогенез наследственной гипертензии с гиперкалиемией: ингибируется котранспортер Na-Cl диким типом, но не мутантным WNK4.Proc Natl Acad Sci USA 100(2):680–684. дои: 10.1073/pnas.242735399
КАС пабмед ПабМед Центральный Статья Google Scholar
Yang CL, Angell J, Mitchell R, Ellison DH (2003)Киназы Wnk регулируют тиазид-чувствительный котранспорт Na-Cl.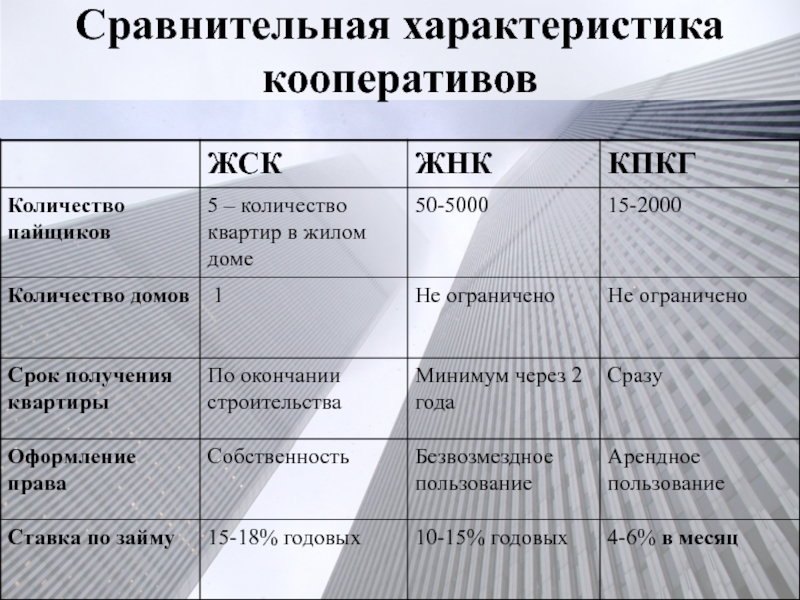 J Clin Invest 111 (7): 1039–1045. дои: 10.1172/jci17443
J Clin Invest 111 (7): 1039–1045. дои: 10.1172/jci17443
КАС пабмед ПабМед Центральный Статья Google Scholar
Vitari AC, Deak M, Morrice NA, Alessi DR (2005)Протеинкиназы WNK1 и WNK4, мутировавшие при синдроме гипертензии Гордона, фосфорилируют и активируют протеинкиназы SPAK и OSR1. Biochem J 391 (Pt 1): 17–24. дои: 10.1042/BJ20051180
КАС пабмед ПабМед Центральный Статья Google Scholar
Gagnon KB, Delpire E (2012)Молекулярная физиология SPAK и OSR1: две родственные ste20 протеинкиназы, регулирующие транспорт ионов.Physiol Rev 92 (4): 1577–1617. doi:10.1152/physrev.00009.2012
КАС пабмед ПабМед Центральный Статья Google Scholar
Piechotta K, Garbarini N, England R, Delpire E (2003) Характеристика взаимодействия стресс-киназы SPAK с котранспортером Na+-K+-2Cl- в нервной системе: доказательства опорной роли киназа. J Biol Chem 278(52):52848–52856. дои: 10.1074/jbc.М309436200
J Biol Chem 278(52):52848–52856. дои: 10.1074/jbc.М309436200
КАС пабмед Статья Google Scholar
Рафики Ф.Х., Зубер А.М., Гловер М., Ричардсон С., Флеминг С., Йованович С., Йованович А., О’Шонесси К.М., Алесси Д.Р. (2010)Роль WNK-активируемой киназы SPAK в регулировании артериального давления. EMBO Mol Med 2 (2): 63–75. дои: 10.1002/emmm.2008
КАС пабмед ПабМед Центральный Статья Google Scholar
McCormick JA, Mutig K, Nelson JH, Saritas T, Hoorn EJ, Yang CL, Rogers S, Curry J, Delpire E, Bachmann S, Ellison DH (2011)Изоформный переключатель SPAK модулирует почечный транспорт соли и кровяное давление. Cell Metab 14 (3): 352–364. doi:10.1016/j.cmet.2011.07.009
КАС пабмед ПабМед Центральный Статья Google Scholar

Rinehart J, Vazquez N, Kahle KT, Hodson CA, Ring AM, Gulcicek EE, Louvi A, Bobadilla NA, Gamba G, Lifton RP (2011) Киназа WNK2 является новым регулятором основного нейронального катион-хлорида котранспортеры.J Biol Chem 286(34):30171–30180. дои: 10.1074/jbc.M111.222893
КАС пабмед ПабМед Центральный Статья Google Scholar
Filippi BM, de los Heros P, Mehellou Y, Navratilova I, Gourlay R, Deak M, Plater L, Toth R, Zeqiraj E, Alessi DR (2011) Mo25 является главным регулятором SPAK/OSR1 и Протеинкиназы MST3/MST4/YSK1. EMBO J 30 (9): 1730–1741. doi: 10.1038 / emboj.2011.78
Vitari AC, Thastrup J, Rafiqi FH, Deak M, Morrice NA, Karlsson HK, Alessi DR (2006) Функциональные взаимодействия киназ SPAK/OSR1 с их вышестоящим активатором WNK1 и нижележащим субстратом NKCC1. Биохим J 397 (1): 223–231. дои: 10.1042/bj20060220
КАС пабмед ПабМед Центральный Статья Google Scholar

Villa F, Goebel J, Rafiqi FH, Deak M, Thastrup J, Alessi DR, van Aalten DM (2007) Структурное понимание распознавания субстратов и активаторов киназой OSR1.EMBO Rep 8 (9): 839–845. doi:10.1038/sj.embor.7401048
КАС пабмед ПабМед Центральный Статья Google Scholar
Zhang J, Siew K, Macartney T, O’Shaughnessy KM, Alessi DR (2015) Критическая роль домена CCT протеинкиназы SPAK в контроле артериального давления. Хум Мол Генет 24 (16): 4545–4558. дои: 10.1093/hmg/ddv185
КАС пабмед ПабМед Центральный Статья Google Scholar
Richardson C, Rafiqi FH, Karlsson HK, Moleleki N, Vandewalle A, Campbell DG, Morrice NA, Alessi DR (2008) Активация тиазид-чувствительного котранспортера Na+–Cl– регулируемыми WNK киназами SPAK и OSR1. J Cell Sci 121 (Pt 5): 675–684. дои: 10.1242/jcs.025312
КАС пабмед Статья Google Scholar

Yang SS, Lo YF, Wu CC, Lin SW, Yeh CJ, Chu P, Sytwu HK, Uchida S, Sasaki S, Lin SH (2010) Мыши с нокаутом SPAK проявляют синдром Гительмана и нарушение вазоконстрикции.J Am Soc Nephrol 21 (11): 1868–1877. doi: 10.1681 / asn.200
95КАС пабмед ПабМед Центральный Статья Google Scholar
Lin SH, Yu IS, Jiang ST, Lin SW, Chu P, Chen A, Sytwu HK, Sohara E, Uchida S, Sasaki S, Yang SS (2011) Нарушение фосфорилирования Na(+)–K (+)–2Cl(–) котранспортера при дефиците киназы-1, чувствительной к окислительному стрессу, проявляется гипотонией и синдромом Барттера.Proc Natl Acad Sci USA 108(42):17538–17543. doi:10.1073/pnas.1107452108
КАС пабмед ПабМед Центральный Статья Google Scholar
Xie J, Yoon J, Yang SS, Lin SH, Huang CL (2013) Протеинкиназа WNK1 регулирует эмбриональное сердечно-сосудистое развитие посредством сигнального каскада OSR1. J Biol Chem 288(12):8566–8574. дои: 10.1074/jbc.M113.451575
J Biol Chem 288(12):8566–8574. дои: 10.1074/jbc.M113.451575
КАС пабмед ПабМед Центральный Статья Google Scholar
Ferdaus MZ, Barber KW, Lopez-Cayuqueo KI, Terker AS, Argaiz ER, Gassaway BM, Chambrey R, Gamba G, Rinehart J, McCormick JA (2016) SPAK и OSR1 играют важную роль в гомеостазе калия, воздействуя на дистальные извилистые трубочка. J Physiol 594 (17): 4945–4966. дои: 10.1113/jp272311
КАС пабмед Статья Google Scholar
Траганте В., Барнс Майкл Р., Ганеш Санти К., Ланктри Мэтью Б., Го В., Франческини Н., Смит Эрин Н., Джонсон Т., Холмс Майкл В., Падманабхан С., Карчевски Конрад Дж., Альмогера Б., Барнард Дж. , Баумерт Дж., Чанг И-Пей С., Элберс Клара С., Фаррал М., Фишер Мэри Э., Гаунт Том Р., Го Йоханнес М., Гигер С., Гоэл А., Гонг Ю., Исаакс А., Клебер Маркус Э., Лич Айрин М. , Макдонаф Кейтрин В., Мейс Маттейс Ф., Меландер О., Нельсон Кристофер П., Нолте Илья М., Панкрац Н., Прайс Том С., Шаффер Дж., Шах С., Томашевски М., ван дер Мост Питер Дж., Ван Иперен Эрик П., Вонк Джудит М., Витковска К., Вонг Кэролайн О., Чжан Л., Бейтелшис Эмбер Л., Беренсон Джеральд С., Бхатт Дипак Л., Браун М., Берт А., Купер-ДеХофф Ронда М., Коннелл Джон М., Круикшенкс Карен Дж., Кертис Шон П., Дэви-Смит Г., Деллес С., Гансевурт Рон Т., Го Х, Хайцин С., Хасти Клэр Э., Хофкер Мартен Х., Ховинг Г.К., Ким Дэниел С., Киркланд Сьюзен А., Клейн Барбара Э., Клейн Р., Ли Юн Р., Майвальд С., Ньютон-Чех С, О’Брайен Эоин Т, Он Лэнд-Море Н.К., Палмас В., Парса А., Пеннинкс Бренда В., Петтингер М., Васан Р.С., Ранчалис Джейн Э., М. Ридкер П., Роуз Линда М., Север П., Шимбо Д., Стил Л., Столк Рональд П., Торанд Б., Трип Мике Д., ван Дуйн Корнелия М., Вершурен В.М., Вийменга С., Вятт С., Янг Дж. Х., Звиндерман Эйлко Х., Беззина Конни Р., Бурвинкль Э., Касас Хуан П., Колфилд Марк Дж., Чакраварти А.
, Макдонаф Кейтрин В., Мейс Маттейс Ф., Меландер О., Нельсон Кристофер П., Нолте Илья М., Панкрац Н., Прайс Том С., Шаффер Дж., Шах С., Томашевски М., ван дер Мост Питер Дж., Ван Иперен Эрик П., Вонк Джудит М., Витковска К., Вонг Кэролайн О., Чжан Л., Бейтелшис Эмбер Л., Беренсон Джеральд С., Бхатт Дипак Л., Браун М., Берт А., Купер-ДеХофф Ронда М., Коннелл Джон М., Круикшенкс Карен Дж., Кертис Шон П., Дэви-Смит Г., Деллес С., Гансевурт Рон Т., Го Х, Хайцин С., Хасти Клэр Э., Хофкер Мартен Х., Ховинг Г.К., Ким Дэниел С., Киркланд Сьюзен А., Клейн Барбара Э., Клейн Р., Ли Юн Р., Майвальд С., Ньютон-Чех С, О’Брайен Эоин Т, Он Лэнд-Море Н.К., Палмас В., Парса А., Пеннинкс Бренда В., Петтингер М., Васан Р.С., Ранчалис Джейн Э., М. Ридкер П., Роуз Линда М., Север П., Шимбо Д., Стил Л., Столк Рональд П., Торанд Б., Трип Мике Д., ван Дуйн Корнелия М., Вершурен В.М., Вийменга С., Вятт С., Янг Дж. Х., Звиндерман Эйлко Х., Беззина Конни Р., Бурвинкль Э., Касас Хуан П., Колфилд Марк Дж., Чакраварти А. , Часман Дэниел И., Дэвидсон Карина В., Довенданс Питер А., Доминикак Анна Ф., Фитцджеральд Гаррет А., Гамс Джон Г., Форнейдж М., Хаконарсон Х., Гальдер И., Хиллеге Ханс Л., Иллиг Т., Ярвик Гейл П., Джонсон Джули А., Кастелейн Джон Дж., Кениг В., Кумари М., Марц В., Мюррей Сара С., О’Коннелл Джеффри Р., Олдехинкель Альбертин Дж., Панков Джеймс С., Рейдер Даниэль Дж., Редлайн С., Рейли Муредах П., Шадт Эрик Э., Коттке-Марчант К., Снидер Х., Снайдер М., Стэнтон Элис В., Тобин Мартин Д., Уиттерлинден Андре Г., ван дер Харст П., ван дер Шоу Ивонн Т., Самани Нилеш Дж., Уоткинс Х., Джонсон Эндрю Д., Райнер Алекс П., Чжу Х, де Баккер Пол И., Леви Д., Ассельбергс Фолкерт В, Манро Патриция Б., Китинг Брендан Дж. (2014) Геноцентрический метаанализ 87 736 лиц европейского происхождения выявил несколько локусов, связанных с артериальным давлением.Am J Hum Genet 94 (3): 349–360. doi: 10.1016 / j.ajhg.2013.12.016
, Часман Дэниел И., Дэвидсон Карина В., Довенданс Питер А., Доминикак Анна Ф., Фитцджеральд Гаррет А., Гамс Джон Г., Форнейдж М., Хаконарсон Х., Гальдер И., Хиллеге Ханс Л., Иллиг Т., Ярвик Гейл П., Джонсон Джули А., Кастелейн Джон Дж., Кениг В., Кумари М., Марц В., Мюррей Сара С., О’Коннелл Джеффри Р., Олдехинкель Альбертин Дж., Панков Джеймс С., Рейдер Даниэль Дж., Редлайн С., Рейли Муредах П., Шадт Эрик Э., Коттке-Марчант К., Снидер Х., Снайдер М., Стэнтон Элис В., Тобин Мартин Д., Уиттерлинден Андре Г., ван дер Харст П., ван дер Шоу Ивонн Т., Самани Нилеш Дж., Уоткинс Х., Джонсон Эндрю Д., Райнер Алекс П., Чжу Х, де Баккер Пол И., Леви Д., Ассельбергс Фолкерт В, Манро Патриция Б., Китинг Брендан Дж. (2014) Геноцентрический метаанализ 87 736 лиц европейского происхождения выявил несколько локусов, связанных с артериальным давлением.Am J Hum Genet 94 (3): 349–360. doi: 10.1016 / j.ajhg.2013.12.016
Манро П.Б., Барнс М.Р., Колфилд М.Дж. (2013) Достижения в области геномики артериального давления. Циркуляр рез. 112 (10): 1365–1379. doi:10.1161/circresaha.112.300387
Циркуляр рез. 112 (10): 1365–1379. doi:10.1161/circresaha.112.300387
КАС пабмед Статья Google Scholar
Ван Ю., О’Коннелл Дж. Р., МакАрдл П. Ф., Уэйд Дж. Б., Дорфф С. Э., Шах С. Дж., Ши Х., Пан Л., Рамперсо Э., Шен Х., Ким Д. Д., Субраманья А. Р., Штайнле Н. И., Парса А., Ober CC, Welling PA, Chakravarti A, Weder AB, Cooper RS, Mitchell BD, Shuldiner AR, Chang Y-PC (2009) Исследование ассоциации всего генома идентифицирует stk39 как ген предрасположенности к гипертонии.Proc Natl Acad Sci USA 106(1):226–231. doi:10.1073/pnas.0808358106
КАС пабмед Статья Google Scholar
Кидамби С., Гош С., Котчен Дж. М., Грим К. Э., Кришнасвами С., Калдунски М. Л., Коули А. В., Патель С. Б., Котчен Т. А. (2012) Неповторяющееся исследование полногеномного исследования ассоциации гипертонии и крови давление у афроамериканцев. БМС Мед Жене 13:27. дои: 10.1186/1471-2350-13-27
БМС Мед Жене 13:27. дои: 10.1186/1471-2350-13-27
КАС пабмед ПабМед Центральный Статья Google Scholar
Xi B, Zhao X, Chandak GR, Shen Y, Cheng H, Hou D, Wang X, Mi J (2013) Влияние ожирения на связь между генетическими вариантами, выявленными в ходе полногеномных исследований ассоциации, и риском гипертонии у китайских детей. Am J Hypertens 26 (8): 990–996. дои: 10.1093/ajh/hpt046
КАС пабмед Статья Google Scholar
Xi B, Chen M, Chandak GR, Shen Y, Yan L, He J, Mou S-H (2013) Полиморфизм Stk39 связан с гипертонической болезнью: систематический обзор и метаанализ.PLoS One 8(3):e59584. doi:10.1371/journal.pone.0059584
КАС пабмед ПабМед Центральный Статья Google Scholar
Zhao H, Qi Y, Wang Y, Wang Y, Lu C, Xiao Y, Wang B, Niu W (2014) Интерактивный вклад множественных полиморфизмов гена серин/треонинкиназы 39 в гипертонию среди северо-восточных ханьцев. Научный отчет 4:5116. дои: 10.1038/srep05116
Научный отчет 4:5116. дои: 10.1038/srep05116
КАС пабмед ПабМед Центральный Google Scholar
Шао Л., Ланг Ю., Ван Ю., Гао Ю., Чжан В., Ниу Х., Лю С., Чен Н. (2012) Высокочастотный вариант с. T60m в котранспортере NaCl и вариабельность артериального давления у китайцев хань. Am J Nephrol 35 (6): 515–519. дои: 10.1159/000339165
КАС пабмед Статья Google Scholar
Yang SS, Fang YW, Tseng MH, Chu PY, Yu IS, Wu HC, Lin SW, Chau T, Uchida S, Sasaki S, Lin YF, Sytwu HK, Lin SH (2013) Фосфорилирование регулирует NCC стабильность и транспортную активность in vivo.J Am Soc Nephrol 24 (10): 1587–1597. doi: 10.1681 / asn.2012070742
КАС пабмед ПабМед Центральный Статья Google Scholar
Yang CL, Zhu X, Wang Z, Subramanya AR, Ellison DH (2005) Механизмы взаимодействия WNK1 и WNK4 в регуляции тиазид-чувствительного котранспорта nacl. J Clin Invest 115 (5): 1379–1387. дои: 10.1172/jci22452
J Clin Invest 115 (5): 1379–1387. дои: 10.1172/jci22452
КАС пабмед ПабМед Центральный Статья Google Scholar
Эллисон Д.Х. (2003)Тиазид-чувствительный котранспортер Na-Cl и болезни человека: возрождение старого игрока. J Am Soc Nephrol 14(2):538–540
PubMed Google Scholar
Moriguchi T, Urushiyama S, Hisamoto N, Iemura S, Uchida S, Natsume T, Matsumoto K, Shibuya H (2005) WNK1 регулирует фосфорилирование катион-хлорид-связанных котранспортеров через киназы, связанные с ste20, SPAK и ОСР1. J Biol Chem 280(52):42685–42693.дои: 10.1074/jbc.M510042200
КАС пабмед Статья Google Scholar
Xie J, Wu T, Xu K, Huang IK, Cleaver O, Huang C-L (2009) Эндотелиально-специфическая экспрессия киназы WNK1 необходима для ангиогенеза и развития сердца у мышей. Ам Дж. Патол 175 (3): 1315–1327. дои: 10.2353/ajpath.2009.0
Ам Дж. Патол 175 (3): 1315–1327. дои: 10.2353/ajpath.2009.0
КАС пабмед ПабМед Центральный Статья Google Scholar
Zambrowicz BP, Abuin A, Ramirez-Solis R, Richter LJ, Piggott J, BeltrandelRio H, Buxton EC, Edwards J, Finch RA, Friddle CJ, Gupta A, Hansen G, Hu Y, Huang W, Jaing C, Key BW , Кипп П., Кольхауф Б., Ма З.К., Маркесич Д., Пейн Р., Поттер Д.Г., Цянь Н., Шоу Дж., Шрик Дж., Ши З.З., Спаркс М.Дж., Ван Слигтенхорст И., Фогель П., Уоке В., Сюй Н., Чжу К., Person C, Sands AT (2003) Дефицит киназы WNK1 снижает кровяное давление у мышей: скрининг генных ловушек для выявления потенциальных целей для терапевтического вмешательства.Proc Natl Acad Sci 100 (24): 14109–14114. doi:10.1073/pnas.2336103100
ПабМед ПабМед Центральный Статья Google Scholar
Pacheco-Alvarez D, Vazquez N, Castaneda-Bueno M, de-Los-Heros P, Cortes-Gonzalez C, Moreno E, Meade P, Bobadilla NA, Gamba G (2012) Взаимодействие WNK3–SPAK требуется для модуляции NCC и других членов семейства slc12. Cell Physiol Biochem 29 (1–2): 291–302.дои: 10.1159/000337610
Cell Physiol Biochem 29 (1–2): 291–302.дои: 10.1159/000337610
Rinehart J, Kahle KT, de Los Heros P, Vazquez N, Meade P, Wilson FH, Hebert SC, Gimenez I, Gamba G, Lifton RP (2005) Киназа WNK3 является положительным регулятором NKCC2 и NCC , почечный катион-Cl-котранспортер, необходимый для нормального гомеостаза артериального давления. Proc Natl Acad Sci USA 102(46):16777–16782. doi:10.1073/pnas.0508303102
КАС пабмед ПабМед Центральный Статья Google Scholar
Glover M, Zuber AM, O’Shaughnessy KM (2009)Изоформы WNK3 в почках и головном мозге оказывают противоположное влияние на экспрессию NCCT. J Am Soc Nephrol 20 (6): 1314–1322. doi: 10.1681 / asn.2008050542
КАС пабмед ПабМед Центральный Статья Google Scholar
Oi K, Sohara E, Rai T, Misawa M, Chiga M, Alessi DR, Sasaki S, Uchida S (2012) Незначительная роль WNK3 в регуляции фосфорилирования почечных котранспортеров NKCC2 и NCC in vivo . Биол Открытый 1 (2): 120–127. дои: 10.1242/био.2011048
Биол Открытый 1 (2): 120–127. дои: 10.1242/био.2011048
КАС пабмед Статья Google Scholar
Базуа-Валенти С., Чавес-Каналес М., Рохас-Вега Л., Гонсалес-Родригес Х., Васкес Н., Родригес-Гама А., Аргаис Э.Р., Мело З., Плата С., Эллисон Д.Х., Гарсия-Вальдес Х. , Hadchouel J, Gamba G (2015)Влияние WNK4 на котранспортер Na+-Cl- модулируется внутриклеточным хлоридом. J Am Soc Nephrol 26 (8): 1781–1786.doi: 10.1681 / asn.2014050470
ПабМед Статья КАС Google Scholar
Gamba G (2012) Регуляция почечного котранспортера Na+–Cl– путем фосфорилирования и убиквитинирования. Am J Physiol Renal Physiol 303(12):F1573–F1583. doi: 10.1152/ajprenal.00508.2012
КАС пабмед ПабМед Центральный Статья Google Scholar

Castaneda-Bueno M, Cervantes-Perez LG, Vazquez N, Uribe N, Kantesaria S, Morla L, Bobadilla NA, Doucet A, Alessi DR, Gamba G (2012) Активация почечного котранспортера Na+:Cl- ангиотензином II процесс, зависящий от WNK4. Proc Natl Acad Sci USA 109(20):7929–7934. doi:10.1073/pnas.1200947109
КАС пабмед ПабМед Центральный Статья Google Scholar
Лалиоти М.Д., Чжан Дж., Фолькман Х.М., Кале К.Т., Хоффманн К.Е., Тока Х.Р., Нельсон-Уильямс С., Эллисон Д.Х., Флавелл Р., Бут С.Дж., Лу И., Геллер Д.С., Лифтон Р.П. (2006) WNK4 контролирует кровяное давление и гомеостаз калия посредством регуляции массы и активности дистального извитого канальца.Нат Жене 38 (10): 1124–1132. дои: 10.1038/ng1877
КАС пабмед Статья Google Scholar
Теркер А.С., Чжан С., Эрспамер К.Дж., Гамба Г., Ян С.Л., Эллисон Д. Х. (2016) Уникальные хлорид-чувствительные свойства WNK4 позволяют дистальным нефронам модулировать гомеостаз калия. Kidney Int 89 (1): 127–134. дои: 10.1038 / ki.2015.289
Х. (2016) Уникальные хлорид-чувствительные свойства WNK4 позволяют дистальным нефронам модулировать гомеостаз калия. Kidney Int 89 (1): 127–134. дои: 10.1038 / ki.2015.289
КАС пабмед ПабМед Центральный Статья Google Scholar
Penton D, Czogalla J, Wengi A, Himmerkus N, Loffing-Cueni D, Carrel M, Rajaram RD, Staub O, Bleich M, Schweda F, Loffing J (2016) Внеклеточный K + быстро контролирует фосфорилирование NCC в нативном DCT с помощью Cl- -зависимые и -независимые механизмы. Дж. Физиол. дои: 10.1113/jp272504
ПабМед Google Scholar
Weinstein AM (2005) Математическая модель дистального извитого канальца крысы. I. Котранспортерная функция в ранней DCT.Am J Physiol Renal Physiol 289(4):F699–F720. doi:10.1152/ajprenal.00043.2005
КАС пабмед Статья Google Scholar

Hadchouel J, Delaloy C, Faure S, Achard JM, Jeunemaitre X (2006) Семейная гиперкалиемическая гипертензия. J Am Soc Nephrol 17 (1): 208–217. doi: 10.1681 / asn.2005030314
КАС пабмед Статья Google Scholar
Genschik P, Sumara I, Lechner E (2013)Новое семейство убиквитинлигаз с кольцом cullin3 (crl3s): клеточные функции и последствия заболеваний. EMBO J 32 (17): 2307–2320. doi: 10.1038 / emboj.2013.173
КАС пабмед ПабМед Центральный Статья Google Scholar
Petroski MD, Deshaies RJ (2005) Функция и регуляция убиквитинлигазы кулинового кольца. Nat Rev Mol Cell Biol 6(1):9–20.дои: 10.1038/nrm1547
КАС пабмед Статья Google Scholar
Гершко А., Чехановер А. (1998) Система убиквитина. Annu Rev Biochem 67: 425–479. doi:10.1146/annurev.biochem.67.1.425
Annu Rev Biochem 67: 425–479. doi:10.1146/annurev.biochem.67.1.425
КАС пабмед Статья Google Scholar (2002) Структура убиквитинлигазного комплекса cul1–rbx1–skp1–f boxskp2 scf.Природа 416 (6882): 703–709. дои: 10.1038/416703a
КАС пабмед Статья Google Scholar
Dhanoa BS, Cogliati T, Satish AG, Bruford EA, Friedman JS (2013) Обновление семейства генов, подобных kelch (klhl). Гум Геном 7:13. дои: 10.1186/1479-7364-7-13
Артикул КАС Google Scholar
Вакабаяси М., Мори Т., Исобе К., Сохара Э., Суса К., Араки Ю., Чига М., Кикути Э., Номура Н., Мори Ю., Мацуо Х., Мурата Т., Номура С., Асано Т., Кавагути Х. , Nonoyama S, Rai T, Sasaki S, Uchida S (2013)Нарушение KLHL3-опосредованного убиквитинирования WNK4 вызывает гипертонию у человека. Ячейка Rep 3 (3): 858–868. doi:10.1016/j.celrep.2013.02.024
Ячейка Rep 3 (3): 858–868. doi:10.1016/j.celrep.2013.02.024
КАС пабмед Статья Google Scholar
Shibata S, Zhang J, Puthumana J, Stone KL, Lifton RP (2013) Kelch-like 3 и cullin 3 регулируют электролитный гомеостаз посредством убиквитинирования и деградации WNK4. Proc Natl Acad Sci USA 110(19):7838–7843. doi:10.1073/pnas.1304592110
КАС пабмед ПабМед Центральный Статья Google Scholar
Суса К., Сохара Э., Рай Т., Зения М., Мори Ю., Мори Т., Чига М., Номура Н., Нисида Х., Такахаши Д., Исобе К., Иноуэ Ю., Такеиси К., Такеда Н., Сасаки С., Учида С. (2014) ) Нарушение деградации киназ WNK1 и WNK4 вызывает PHAII у мышей с нокаутом мутантного KLHL3. Хум Мол Генет 23 (19): 5052–5060. дои: 10.1093/hmg/ddu217
КАС пабмед Статья Google Scholar

Osawa M, Ogura Y, Isobe K, Uchida S, Nonoyama S, Kawaguchi H (2013) Анализ гена Cul3 позволяет проводить раннее вмешательство при педиатрическом псевдогипоальдостеронизме II типа в младенчестве.Педиатр Нефрол 28 (9): 1881–1884. дои: 10.1007/s00467-013-2496-6
ПабМед Статья Google Scholar
Цудзи С., Ямашита М., Униши Г., Такева Р., Кимата Т., Исобе К., Чига М., Учида С., Канеко К. (2013) Маленький ребенок с псевдогипоальдостеронизмом II типа в результате мутации куллина 3. BMC Нефрол 14:166. дои: 10.1186/1471-2369-14-166
КАС пабмед ПабМед Центральный Статья Google Scholar
Маккормик Дж.А., Ян С.Л., Чжан С., Дэвидж Б., Бланкенштейн К.И., Теркер А.С., Ярбро Б., Меермайер Н.П., Парк Х.Дж., Маккалли Б., Уэст М., Борщевски А., Химмеркус Н., Блейх М., Бахманн С., Мутиг К., Аргаиз ER, Gamba G, Singer JD, Ellison DH (2014)Кулин 3, связанный с гиперкалиемической гипертензией, способствует передаче сигналов WNK путем деградации KLHL3. J Clin Invest 124 (11): 4723–4736. дои: 10.1172/JCI76126
J Clin Invest 124 (11): 4723–4736. дои: 10.1172/JCI76126
КАС пабмед ПабМед Центральный Статья Google Scholar
Yang SS, Morimoto T, Rai T, Chiga M, Sohara E, Ohno M, Uchida K, Lin SH, Moriguchi T, Shibuya H, Kondo Y, Sasaki S, Uchida S (2007) Молекулярный патогенез псевдогипоальдостеронизма типа II: поколение и анализ мышиной модели нокина WNK4(d561a/+). Cell Metab 5 (5): 331–344. doi:10.1016/j.cmet.2007.03.009
КАС пабмед Статья Google Scholar
Chowdhury JA, Liu CH, Zuber AM, O’Shaughnessy KM (2013)Индуцибельная модель трансгенных мышей для семейной гипертензии с гиперкалиемией (синдром Гордона или псевдогипоальдостеронизм II типа).Clin Sci (Лондон) 124 (12): 701–708. дои: 10.1042/cs20120430
КАС Статья Google Scholar

Mori T, Kikuchi E, Watanabe Y, Fujii S, Ishigami-Yuasa M, Kagechika H, Sohara E, Rai T, Sasaki S, Uchida S (2013) Скрининг химической библиотеки на ингибиторы передачи сигналов WNK с использованием флуоресцентной корреляции спектроскопия. Biochem J 455 (3): 339–345. дои: 10.1042/bj20130597
КАС пабмед Статья Google Scholar
Barroso I, Gurnell M, Crowley VE, Agostini M, Schwabe JW, Soos MA, Maslen GL, Williams TD, Lewis H, Schafer AJ, Chatterjee VK, O’Rahilly S (1999) Доминантные негативные мутации в PPARgamma человека, связанные с тяжелым инсулинорезистентность, сахарный диабет и артериальная гипертензия. Природа 402 (6764): 880–883. Дои: 10.1038/47254
КАС пабмед Google Scholar
Pelham CJ, Ketsawatsomkron P, Groh S, Grobe JL, de Lange WJ, Ibeawuchi SR, Keen HL, Weatherford ET, Faraci FM, Sigmund CD (2012) Cullin-3 регулирует функцию гладких мышц сосудов и артериальную кровь давление через PPARgamma и RhoA/Rho-киназу. Cell Metab 16 (4): 462–472. doi:10.1016/j.cmet.2012.08.011
Cell Metab 16 (4): 462–472. doi:10.1016/j.cmet.2012.08.011
КАС пабмед ПабМед Центральный Статья Google Scholar
Араки Ю., Рай Т., Сохара Э., Мори Т., Иноуэ Ю., Исобе К., Кикучи Э., Охта А., Сасаки С., Учида С. (2015) Генерация и анализ нокаутированных мышей, несущих псевдогипоальдостеронизм II типа — вызывает мутации в гене cullin 3. Биол Открытый 4 (11): 1509–1517. дои: 10.1242/био.013276
ПабМед ПабМед Центральный Статья Google Scholar
Ibeawuchi SR, Agbor LN, Quelle FW, Sigmund CD (2015)Вызывающие гипертензию мутации в белке cullin3 нарушают убиквитинирование белка RhoA и усиливают связь с адаптерами субстрата. J Biol Chem 290(31):19208–19217. дои: 10.1074/jbc.M115.645358
КАС пабмед ПабМед Центральный Статья Google Scholar

Bergaya S, Faure S, Baudrie V, Rio M, Escoubet B, Bonnin P, Henrion D, Loirand G, Achard JM, Jeunemaitre X, Hadchouel J (2011) WNK1 регулирует сужение сосудов и реакцию артериального давления на α1 -адренергическая стимуляция у мышей.Гипертония 58 (3): 439–445. doi:10.1161/гипертония.111.172429
КАС пабмед Статья Google Scholar (2013) Потребление соли с пищей регулирует каскад фосфорилирования WNK3-SPAK-NKCC1 в аорте мыши через ангиотензин II. Гипертония 62 (5): 872–878. дои: 10.1161/гипертонияга.113.01543
КАС пабмед Статья Google Scholar
Ko B, Mistry A, Hanson L, Mallick R, Hoover RS (2015) Механизмы стимуляции ангиотензином II NCC зависят от времени в клетках MDCT15. Am J Physiol Renal Physiol 308(7):F720–F727. doi: 10.1152/ajprenal.00465.2014
КАС пабмед ПабМед Центральный Статья Google Scholar

Subramanya AR, Liu J, Ellison DH, Wade JB, Welling PA (2009) WNK4 направляет тиазид-чувствительный котранспортер nacl к лизосоме и стимулирует взаимодействие AP-3. J Biol Chem 284(27):18471–18480. дои: 10.1074/jbc.M109.008185
КАС пабмед ПабМед Центральный Статья Google Scholar
Yang CL, Liu X, Paliege A, Zhu X, Bachmann S, Dawson DC, Ellison DH (2007) WNK1 и WNK4 модулируют активность CFTR.Biochem Biophys Res Commun 353 (3): 535–540. doi: 10.1016/j.bbrc.2006.11.151
КАС пабмед Статья Google Scholar
Mendes AI, Matos P, Moniz S, Luz S, Amaral MD, Farinha CM, Jordan P (2011)Антагонистическая регуляция экспрессии трансмембранного регулятора проводимости кистозного фиброза на клеточной поверхности с помощью протеинкиназ WNK4 и тирозинкиназы селезенки. Mol Cell Biol 31(19):4076–4086. дои: 10.1128/мкб.05152-11
дои: 10.1128/мкб.05152-11
КАС пабмед ПабМед Центральный Статья Google Scholar
Luz S, Kongsuphol P, Mendes AI, Romeiras F, Sousa M, Schreiber R, Matos P, Jordan P, Mehta A, Amaral MD, Kunzelmann K, Farinha CM (2011) Вклад казеинкиназы 2 и тирозинкиназа селезенки к переносу CFTR и активности, индуцированной протеинкиназой А. Mol Cell Biol 31(22):4392–4404. doi: 10.1128/mcb.05517-11
КАС пабмед ПабМед Центральный Статья Google Scholar
Lieberman J, Rodbard S (1975) Низкое кровяное давление у молодых людей с муковисцидозом: эффект хронической потери соли с потом? Ann Intern Med 82(6):806–808
CAS пабмед Статья Google Scholar
Peotta VA, Bhandary P, Ogu U, Volk KA, Roghair RD (2014)Снижение артериального давления у носителей CFTR-F508del коррелирует со сниженной артериальной реактивностью, а не с объемом циркулирующей крови у мышей. PLoS One 9(5):e96756.doi:10.1371/journal.pone.0096756
PLoS One 9(5):e96756.doi:10.1371/journal.pone.0096756
ПабМед ПабМед Центральный Статья КАС Google Scholar
Lu C, Pribanic S, Debonneville A, Jiang C, Rotin D (2007) Мотив PY ENaC, мутировавший при синдроме Лиддла, регулирует интернализацию каналов, сортировку и мобилизацию из субапикального пула. Трафик 8 (9): 1246–1264. doi:10.1111/j.1600-0854.2007.00602.x
КАС пабмед Статья Google Scholar
Náray-Fejes-Tóth A, Canessa C, Cleaveland ES, Aldrich G, Fejes-Tóth G (1999) SGK представляет собой альдостерон-индуцируемую киназу в собирательных трубочках почек: воздействие на эпителиальные Na+ каналы. J Biol Chem 274(24):16973–16978. дои: 10.1074/jbc.274.24.16973
ПабМед Статья Google Scholar

Debonneville C, Flores SY, Kamynina E, Plant PJ, Tauxe C, Thomas MA, Münster C, Chraïbi A, Pratt JH, Horisberger JD, Pearce D, Loffing J, Staub O (2001) Фосфорилирование NEDD4 -2 с помощью SGK1 регулирует экспрессию эпителиальных Na+ каналов на клеточной поверхности.EMBO J 20 (24): 7052–7059. doi: 10.1093/emboj/20.24.7052
КАС пабмед ПабМед Центральный Статья Google Scholar
Chen SY, Bhargava A, Mastroberardino L, Meijer OC, Wang J, Buse P, Firestone GL, Verrey F, Pearce D (1999)Эпителиальный натриевый канал, регулируемый альдостерон-индуцированным белком SGK. Proc Natl Acad Sci USA 96(5):2514–2519
CAS пабмед ПабМед Центральный Статья Google Scholar
B-e Xu, Stippec S, Chu P-Y, Lazrak A, Li X-J, Lee BH, English JM, Ortega B, Huang C-L, Cobb MH (2005) WNK1 активирует SGK1 для регуляции эпителиального натриевого канала. Proc Natl Acad Sci USA 102(29):10315–10320. doi:10.1073/pnas.0504422102
Proc Natl Acad Sci USA 102(29):10315–10320. doi:10.1073/pnas.0504422102
Артикул КАС Google Scholar
Heise CJ, Be Xu, Deaton SL, Cha SK, Cheng CJ, Earnest S, Sengupta S, Juan YC, Stippec S, Xu Y, Zhao Y, Huang CL, Cobb MH (2010) Сыворотка и глюкокортикоид -Индуцированная киназа (SGK) 1 и эпителиальный натриевый канал регулируются несколькими членами семейства без лизина (WNK).J Biol Chem 285(33):25161–25167. дои: 10.1074/jbc.M110.103432
КАС пабмед ПабМед Центральный Статья Google Scholar
Yu L, Cai H, Yue Q, Alli AA, Wang D, Al-Khalili O, Bao HF, Eaton DC (2013) Ингибирование ENaC WNK4 не зависит от NEDD4-2-опосредованного убиквитинирования ENaC. Am J Physiol Renal Physiol 305(1):F31–F41. doi: 10.1152/ajprenal.00652.2012
КАС пабмед ПабМед Центральный Статья Google Scholar

Ring AM, Leng Q, Rinehart J, Wilson FH, Kahle KT, Hebert SC, Lifton RP (2007) Сайт SGK1 в WNK4 регулирует активность каналов Na+ и K+ и имеет значение для передачи сигналов альдостерона и гомеостаза K+. Proc Natl Acad Sci 104 (10): 4025–4029. doi:10.1073/pnas.0611728104
КАС пабмед ПабМед Центральный Статья Google Scholar
He G, Wang HR, Huang SK, Huang CL (2007) Intersectin связывает киназы WNK с эндоцитозом ROMK1.J Clin Invest 117 (4): 1078–1087. дои: 10.1172/jci30087
КАС пабмед ПабМед Центральный Статья Google Scholar
Cope G, Murthy M, Golbang AP, Hamad A, Liu CH, Cuthbert AW, O’Shaughnessy KM (2006) WNK1 влияет на поверхностную экспрессию калиевого канала ROMK независимо от WNK4. J Am Soc Nephrol 17 (7): 1867–1874. doi: 10.1681 / asn.2005111224
КАС пабмед Статья Google Scholar

O’Donnell ME (1993) Роль котранспорта Na-K-Cl в регуляции объема эндотелиальных клеток сосудов. Am J Physiol 264 (5 часть 1): C1316–C1326
PubMed Google Scholar
Чипперфилд А.Р., Харпер А.А. (2000) Хлорид в гладких мышцах. Прог Биофиз Мол Биол 74 (3–5): 175–221. дои: 10.1016/S0079-6107(00)00024-9
КАС пабмед Статья Google Scholar
Meyer JW, Flagella M, Sutliff RL, Lorenz JN, Nieman ML, Weber CS, Paul RJ, Shull GE (2002)Снижение артериального давления и тонуса гладкой мускулатуры сосудов у мышей, лишенных базолатерального Na+-K+-2Cl- котранспортера. Am J Physiol Heart Circ Physiol 283 (5): h2846–h2855. doi:10.1152/ajpheart.00083.2002
КАС пабмед Статья Google Scholar
Garg P, Martin CF, Elms SC, Gordon FJ, Wall SM, Garland CJ, Sutliff RL, O’Neill WC (2007) Влияние котранспортера Na-K-2Cl NKCC1 на системное кровяное давление и гладкость мышечный тонус. Am J Physiol Heart Circ Physiol 292 (5): h3100–h3105. doi:10.1152/ajpheart.01402.2006
Am J Physiol Heart Circ Physiol 292 (5): h3100–h3105. doi:10.1152/ajpheart.01402.2006
КАС пабмед ПабМед Центральный Статья Google Scholar
Lai JG, Tsai SM, Tu HC, Chen WC, Kou FJ, Lu JW, Wang HD, Huang CL, Yuh CH (2014) Zebrafish wnk лизин-дефицитная протеинкиназа 1 ( wnk1 ) влияет ангиогенез, связанный с передачей сигналов VEGF. PLoS One 9(8):e106129.doi:10.1371/journal.pone.0106129
ПабМед ПабМед Центральный Статья Google Scholar
Vitari AC, Deak M, Collins BJ, Morrice N, Prescott AR, Phelan A, Humphreys S, Alessi DR (2004) WNK1, киназа, мутировавшая при наследственном синдроме высокого кровяного давления, является новой Субстрат PKB (протеинкиназа B)/AKT. Biochem J 378 (1): 257–268. дои: 10.1042/bj20031692
КАС пабмед ПабМед Центральный Статья Google Scholar

Jiang ZY, Zhou QL, Holik J, Patel S, Leszyk J, Coleman K, Chouinard M, Czech MP (2005) Идентификация WNK1 в качестве субстрата AKT/протеинкиназы B и отрицательного регулятора инсулин-стимулированного митогенеза в 3T3 -клетки L1. J Biol Chem 280(22):21622–21628. дои: 10.1074/jbc.M414464200
КАС пабмед Статья Google Scholar
Nishida H, Sohara E, Nomura N, Chiga M, Alessi DR, Rai T, Sasaki S, Uchida S (2012) Сигнальный путь фосфатидилинозитол-3-киназы/AKT активирует фосфорилирование WNK-OSR1/SPAK-NCC каскад у мышей db/db с гиперинсулинемией.Гипертония 60(4):981–990. doi:10.1161/гипертония.112.201509
КАС пабмед ПабМед Центральный Статья Google Scholar
Small HY, Montezano AC, Rios FJ, Savoia C, Touyz RM (2014)Гипертензия, вызванная антиангиогенной терапией рака ингибиторами фактора роста эндотелия сосудов: понимание и лечение нового синдрома. Can J Cardiol 30 (5): 534–543. doi: 10.1016/j.cjca.2014.02.011
Can J Cardiol 30 (5): 534–543. doi: 10.1016/j.cjca.2014.02.011
ПабМед Статья Google Scholar
Qi W-X, He AN, Shen Z, Yao Y (2013)Заболеваемость и риск гипертонии при применении нового многоцелевого ингибитора киназы акситиниба у онкологических больных: систематический обзор и метаанализ. Br J Clin Pharmacol 76 (3): 348–357. дои: 10.1111/bcp.12149
КАС пабмед ПабМед Центральный Статья Google Scholar
Kikuchi E, Mori T, Zeniya M, Isobe K, Ishigami-Yuasa M, Fujii S, Kagechika H, Ishihara T, Mizushima T, Sasaki S, Sohara E, Rai T, Uchida S (2015) Открытие новых ингибиторов SPAK, которые блокировать передачу сигналов киназы WNK транспортерам катионных хлоридов. J Am Soc Nephrol 26 (7): 1525–1536. doi: 10.1681 / asn.2014060560
КАС пабмед Статья Google Scholar

Ямада К., Парк Х.М., Ригель Д.Ф., ДиПетрилло К., Уэлен Э.Дж., Анисович А., Бейл М., Берстлер Дж., Броклхерст К.Э., Бердик Д.А., Каплан С.Л., Каппарелли М.П., Чен Г., Чен В., Дейл Б. , Дэн Л., Фу Ф., Хамамацу Н., Харасаки К., Герр Т., Хоффманн П., Ху К.Ю., Хуан В.Дж., Идамаканти Н., Имасэ Х., Иваки Ю., Джайн М., Джеясилан Дж., Като М., Кошик В.К., Колс Д., Кунджатхур V, LaSala D, Lee J, Liu J, Luo Y, Ma F, Mo R, Mowbray S, Mogi M, Ossola F, Pandey P, Patel SJ, Raghavan S, Salem B, Shanado YH, Trakshel GM, Turner G, Wakai H, Wang C, Weldon S, Wielicki JB, Xie X, Xu L, Yagi YI, Yasoshima K, Yin J, Yowe D, Zhang JH, Zheng G, Monovich L (2016) Ингибирование низкомолекулярных WNK регулирует сердечно-сосудистую и почечную функция.Nat Chem Biol Adv Online Publ. doi: 10.1038/nchembio.2168. http://www.nature.com/nchembio/journal/vaop/ncurrent/abs/nchembio.2168.html#дополнительная информация
Саритас Т., Борщевски А., Маккормик Дж. А., Палиеж А. , Дате К., Учида С., Теркер А., Химмеркус Н., Блейх М., Демарец С., Лагмани К., Дельпир Э., Эллисон Д. Х., Бахманн С., Мутиг К. (2013) Spak по-разному опосредует эффекты вазопрессина на котранспортеры натрия. J Am Soc Nephrol 24(3):407–418. дои: 10.1681/асн.2012040404
, Дате К., Учида С., Теркер А., Химмеркус Н., Блейх М., Демарец С., Лагмани К., Дельпир Э., Эллисон Д. Х., Бахманн С., Мутиг К. (2013) Spak по-разному опосредует эффекты вазопрессина на котранспортеры натрия. J Am Soc Nephrol 24(3):407–418. дои: 10.1681/асн.2012040404
КАС пабмед ПабМед Центральный Статья Google Scholar
Cheng CJ, Yoon J, Baum M, Huang CL (2015) Ste20/sps1-родственная пролин/богатая аланином киназа (SPAK) имеет решающее значение для реабсорбции натрия в изолированной перфузируемой толстой восходящей конечности. Am J Physiol Renal Physiol 308(5):F437–F443. doi: 10.1152/ajprenal.00493.2013
КАС пабмед Статья Google Scholar
Grimm PR, Lazo-Fernandez Y, Delpire E, Wall SM, Dorsey SG, Weinman EJ, Coleman R, Wade JB, Welling PA (2015) Интегрированная компенсаторная сеть активируется в отсутствие фосфорилирования NCC. J Clin Invest 125 (5): 2136–2150. дои: 10.1172/jci78558
J Clin Invest 125 (5): 2136–2150. дои: 10.1172/jci78558
ПабМед ПабМед Центральный Статья Google Scholar
Клетки | Бесплатный полнотекстовый | Рецептор, чувствительный к кальцию, и регуляция киназ WNK в почках
Гомеостаз кальция имеет решающее значение для нормального сердечного ритма и функций нейронов.Уровни кальция в сыворотке поддерживаются в узком диапазоне 2,2–2,6 ммоль/л благодаря согласованной работе многих органов. Почки в основном участвуют в этом процессе, регулируя экскрецию кальция в соответствии с потребностями организма. Для выполнения этой задачи почечный транспортирующий эпителий снабжен рецепторами к паратиреоидному гормону (ПТГ), кальцитриолу и кальцитонину, обеспечивающим адаптивные реакции на системные сдвиги кальциевого гомеостаза [1,2,3,4]. Эти эндокринные механизмы дополняются способностью почечного эпителия непосредственно воспринимать кальций сыворотки через кальций-чувствительный рецептор (CaSR) [5]. CaSR принадлежит к семейству рецепторов, связанных с G-белком. При связывании Ca 2+ CaSR активирует G-белки G q/11 , G i и G 12/13 , которые стимулируют фосфолипазу C (PLC)-зависимый инозитол-1,4,5-трифосфат. биосинтез с последующим высвобождением Ca 2+ из внутриклеточных депо (обзор см. в [6,7]). Внутриклеточный Ca 2+ [Ca 2+ ] i является важным вторичным мессенджером, влияющим на множество Ca 2+ -чувствительных ферментов, чтобы управлять жизненно важными биологическими событиями клетки, такими как транскрипция генов или везикулярный транспорт [8].С помощью этих механизмов CaSR модулирует экспрессию, содержание на поверхности и фосфорилирование транспортных белков, опосредующих реабсорбцию кальция в почечном эпителии [9,10,11]. Поскольку обработка кальция в дистальном отделе нефрона функционально связана с транспортом натрия, передача сигналов CaSR также участвует в гомеостазе объема и контроле артериального давления [12,13,14,15].
CaSR принадлежит к семейству рецепторов, связанных с G-белком. При связывании Ca 2+ CaSR активирует G-белки G q/11 , G i и G 12/13 , которые стимулируют фосфолипазу C (PLC)-зависимый инозитол-1,4,5-трифосфат. биосинтез с последующим высвобождением Ca 2+ из внутриклеточных депо (обзор см. в [6,7]). Внутриклеточный Ca 2+ [Ca 2+ ] i является важным вторичным мессенджером, влияющим на множество Ca 2+ -чувствительных ферментов, чтобы управлять жизненно важными биологическими событиями клетки, такими как транскрипция генов или везикулярный транспорт [8].С помощью этих механизмов CaSR модулирует экспрессию, содержание на поверхности и фосфорилирование транспортных белков, опосредующих реабсорбцию кальция в почечном эпителии [9,10,11]. Поскольку обработка кальция в дистальном отделе нефрона функционально связана с транспортом натрия, передача сигналов CaSR также участвует в гомеостазе объема и контроле артериального давления [12,13,14,15]. Дистальный отдел нефрона включает толстый восходящий отдел (TAL), дистальный извитой каналец (DCT) и соединительный каналец (CNT). Последующий кортикальный собирательный канал (CD) функционально связан с дистальным отделом нефрона, хотя он происходит из зачатка мочеточника, как и другие части CD, и не принадлежит нефрону с анатомической точки зрения [16,17,18].Появляется все больше доказательств зависимой от CaSR регуляции котранспортера Na + -K + -2Cl — (NKCC2) в TAL и котранспортера Na + -Cl — (NCC) в DCT [12]. ,13] (обзор см. в [19]). NKCC2 имеет решающее значение для концентрации мочи и гомеостаза объема [20,21]. NCC играет ключевую роль в точной настройке экскреции NaCl с мочой и, кроме того, участвует в обработке почек почками [22, 23] (для обзора [24]). Степень реабсорбции NaCl при DCT определяет люминальную нагрузку Na + нижестоящих УНТ и CD, таким образом, являясь этапом, ограничивающим скорость реабсорбции Na + через эпителиальный натриевый канал (ENaC), который электрогенно связан в просвет К + — секреция через наружный мозговой канал почки (ROMK, Kir1.
Дистальный отдел нефрона включает толстый восходящий отдел (TAL), дистальный извитой каналец (DCT) и соединительный каналец (CNT). Последующий кортикальный собирательный канал (CD) функционально связан с дистальным отделом нефрона, хотя он происходит из зачатка мочеточника, как и другие части CD, и не принадлежит нефрону с анатомической точки зрения [16,17,18].Появляется все больше доказательств зависимой от CaSR регуляции котранспортера Na + -K + -2Cl — (NKCC2) в TAL и котранспортера Na + -Cl — (NCC) в DCT [12]. ,13] (обзор см. в [19]). NKCC2 имеет решающее значение для концентрации мочи и гомеостаза объема [20,21]. NCC играет ключевую роль в точной настройке экскреции NaCl с мочой и, кроме того, участвует в обработке почек почками [22, 23] (для обзора [24]). Степень реабсорбции NaCl при DCT определяет люминальную нагрузку Na + нижестоящих УНТ и CD, таким образом, являясь этапом, ограничивающим скорость реабсорбции Na + через эпителиальный натриевый канал (ENaC), который электрогенно связан в просвет К + — секреция через наружный мозговой канал почки (ROMK, Kir1. 1) на этих участках [23] (для обзора [25]). Мутации потери функции в генах, кодирующих NKCC2 или NCC, вызывают гипокалиемические тубулопатии с потерей солей, известные как синдромы Барттера и Гительмана соответственно [26,27]. Точно так же мутации с усилением функции в гене CaSR вызывают потерю солей из-за подавления активности NKCC2 [12,28]. У пациентов с синдромом Барттера, включая его CaSR-зависимую форму, обычно наблюдается активация ренин-ангиотензин-альдостероновой системы (РААС) без сопутствующей прессорной реакции, что можно объяснить снижением чувствительности рецепторов ангиотензина II (AngII) из-за хронического повышения уровня AngII в плазме.Изменение метаболизма простадландинов также может способствовать нарушению сосудистой реактивности у таких пациентов [29]. В качестве альтернативы было показано, что ингибирование передачи сигналов CaSR в почках стимулирует NKCC2 [30]. Чрезмерная реабсорбция соли в дистальном отделе нефрона, опосредованная NKCC2 или NCC, все чаще признается важным патогенетическим фактором солечувствительной гипертензии (для обзора [31,32]).
1) на этих участках [23] (для обзора [25]). Мутации потери функции в генах, кодирующих NKCC2 или NCC, вызывают гипокалиемические тубулопатии с потерей солей, известные как синдромы Барттера и Гительмана соответственно [26,27]. Точно так же мутации с усилением функции в гене CaSR вызывают потерю солей из-за подавления активности NKCC2 [12,28]. У пациентов с синдромом Барттера, включая его CaSR-зависимую форму, обычно наблюдается активация ренин-ангиотензин-альдостероновой системы (РААС) без сопутствующей прессорной реакции, что можно объяснить снижением чувствительности рецепторов ангиотензина II (AngII) из-за хронического повышения уровня AngII в плазме.Изменение метаболизма простадландинов также может способствовать нарушению сосудистой реактивности у таких пациентов [29]. В качестве альтернативы было показано, что ингибирование передачи сигналов CaSR в почках стимулирует NKCC2 [30]. Чрезмерная реабсорбция соли в дистальном отделе нефрона, опосредованная NKCC2 или NCC, все чаще признается важным патогенетическим фактором солечувствительной гипертензии (для обзора [31,32]). Два транспортера принадлежат к семейству электронейтральных катион-связанных котранспортеров хлоридов и разделяют посттрансляционную регуляцию путем фосфорилирования или дефосфорилирования консервативных N-концевых остатков треонина или серина (для обзора [33]).Их активирующее фосфорилирование обеспечивается киназным каскадом, включающим чувствительные к хлориду киназы без лизина [K] (WNK) и две нижестоящие киназы с высокой степенью гомологии, Ste20/SPS1-родственная пролин-аланин-богатая киназа (SPAK) и киназа 1, чувствительная к окислительному стрессу (OSR1) [33]. Мутации с усилением функции в генах, кодирующих WNK1 или WNK4, вызывают семейную гиперкалиемическую гипертензию (FHHt), также известную как псевдогипоальдостеронизм 2 типа или синдром Гордона [34]. Сходным образом нарушение убиквитинирования и деградация WNK из-за мутаций в генах, кодирующих kelch-подобный 3 (KLHL3) или cullin 3, были связаны с FHHt [35].Усиленное фосфорилирование NCC активированным киназным путем WNK-SPAK было признано основным патогенетическим механизмом FHHt, приводящим к задержке соли в почках, увеличению объема и гипертензии [33,34].
Два транспортера принадлежат к семейству электронейтральных катион-связанных котранспортеров хлоридов и разделяют посттрансляционную регуляцию путем фосфорилирования или дефосфорилирования консервативных N-концевых остатков треонина или серина (для обзора [33]).Их активирующее фосфорилирование обеспечивается киназным каскадом, включающим чувствительные к хлориду киназы без лизина [K] (WNK) и две нижестоящие киназы с высокой степенью гомологии, Ste20/SPS1-родственная пролин-аланин-богатая киназа (SPAK) и киназа 1, чувствительная к окислительному стрессу (OSR1) [33]. Мутации с усилением функции в генах, кодирующих WNK1 или WNK4, вызывают семейную гиперкалиемическую гипертензию (FHHt), также известную как псевдогипоальдостеронизм 2 типа или синдром Гордона [34]. Сходным образом нарушение убиквитинирования и деградация WNK из-за мутаций в генах, кодирующих kelch-подобный 3 (KLHL3) или cullin 3, были связаны с FHHt [35].Усиленное фосфорилирование NCC активированным киназным путем WNK-SPAK было признано основным патогенетическим механизмом FHHt, приводящим к задержке соли в почках, увеличению объема и гипертензии [33,34]. Помимо редких моногенетических гипертензивных синдромов, передача сигналов WNK-SPAK-NCC была вовлечена в патофизиологию солечувствительных форм гипертензии, вызванных гиперактивностью РААС или повышенным симпатическим тонусом [36,37]. Дефосфорилирование NKCC2 и NCC опосредуется кальцинейрином, который представляет собой Ca 2+ /кальмодулин (CaM)-зависимую серин/треонинфосфатазу [38,39,40].Кальциневрин подавляет активность WNK-SPAK/OSR1, способствуя их деградации, что снижает количество фосфорилированных NKCC2 и NCC [39,40,41]. Следовательно, функциональные взаимодействия между Cl — -зависимыми и Ca 2+ -зависимыми фосфоферментами определяют степень реабсорбции солей при TAL и DCT. Системные сдвиги уровней внеклеточного кальция [Ca 2+ ] могут влиять на [Ca 2+ ] i в клетках дистального отдела нефрона через активность CaSR, тем самым модулируя перенос почечной соли.В этом контексте ранние эпидемиологические исследования показали повышенную частоту гипертензии у лиц с ограниченным потреблением кальция с пищей (для обзора [42]).
Помимо редких моногенетических гипертензивных синдромов, передача сигналов WNK-SPAK-NCC была вовлечена в патофизиологию солечувствительных форм гипертензии, вызванных гиперактивностью РААС или повышенным симпатическим тонусом [36,37]. Дефосфорилирование NKCC2 и NCC опосредуется кальцинейрином, который представляет собой Ca 2+ /кальмодулин (CaM)-зависимую серин/треонинфосфатазу [38,39,40].Кальциневрин подавляет активность WNK-SPAK/OSR1, способствуя их деградации, что снижает количество фосфорилированных NKCC2 и NCC [39,40,41]. Следовательно, функциональные взаимодействия между Cl — -зависимыми и Ca 2+ -зависимыми фосфоферментами определяют степень реабсорбции солей при TAL и DCT. Системные сдвиги уровней внеклеточного кальция [Ca 2+ ] могут влиять на [Ca 2+ ] i в клетках дистального отдела нефрона через активность CaSR, тем самым модулируя перенос почечной соли.В этом контексте ранние эпидемиологические исследования показали повышенную частоту гипертензии у лиц с ограниченным потреблением кальция с пищей (для обзора [42]). Более того, люди, страдающие солечувствительной гипертензией, по-видимому, особенно восприимчивы к снижению потребления кальция с пищей [43,44,45]. Целью данной обзорной работы является обобщение имеющейся информации о роли CaSR и Ca 2+ -передачи сигналов в WNK-зависимой регуляции дистальной реабсорбции солей.
Более того, люди, страдающие солечувствительной гипертензией, по-видимому, особенно восприимчивы к снижению потребления кальция с пищей [43,44,45]. Целью данной обзорной работы является обобщение имеющейся информации о роли CaSR и Ca 2+ -передачи сигналов в WNK-зависимой регуляции дистальной реабсорбции солей.Последовательность и структурные вариации, определяющие привлечение киназ WNK к лигазе KLHL3 E3
Abstract
Белок BTB-Kelch KLHL3 представляет собой Cullin3-зависимую лигазу E3, которая опосредует убиквитин-зависимую деградацию киназ WNK1-4 для контроля артериального давления и объем клетки. Кристаллическая структура KLHL3 определяет его связывание с кислым мотивом degron, содержащим последовательность PXXP, строго консервативную в WNK1, WNK2 и WNK4. Мутации во втором пролине аннулируют взаимодействие, вызывающее гипертензионный синдром псевдогипоальдостеронизма II типа.WNK3 демонстрирует расходящийся мотив degron, содержащий 4 аминокислотные замены, которые удаляют мотив PXXP, что вызывает вопросы относительно механизма его связывания. Чтобы понять это нетипичное взаимодействие, мы определили кристаллическую структуру домена Келха KLHL3 в комплексе с пептидом WNK3. Электронная плотность позволила впервые проследить полный 11-мерный мотив degron семейства WNK, выявив несколько консервативных особенностей, не учтенных в предыдущей работе, включая дополнительные взаимодействия солевого мостика и водородной связи.В целом, пептид WNK3 принял консервативное положение связывания, за исключением небольшого сдвига, чтобы приспособиться к более объемным аминокислотным заменам на поверхности связывания. В центре второй пролин был заменен на WNK3 Thr541, что дало уникальный фосфорилируемый остаток среди дегронов семейства WNK. Эксперименты по поляризации флуоресценции и структурному моделированию показали, что его фосфорилирование аннулирует взаимодействие KLHL3 аналогично мутациям, вызывающим гипертензию. Вместе эти данные показывают, как домен Kelch KLHL3 может приспосабливаться к связыванию множественных изоформ WNK, и подчеркивают потенциальный регуляторный механизм для рекрутирования WNK3.
Чтобы понять это нетипичное взаимодействие, мы определили кристаллическую структуру домена Келха KLHL3 в комплексе с пептидом WNK3. Электронная плотность позволила впервые проследить полный 11-мерный мотив degron семейства WNK, выявив несколько консервативных особенностей, не учтенных в предыдущей работе, включая дополнительные взаимодействия солевого мостика и водородной связи.В целом, пептид WNK3 принял консервативное положение связывания, за исключением небольшого сдвига, чтобы приспособиться к более объемным аминокислотным заменам на поверхности связывания. В центре второй пролин был заменен на WNK3 Thr541, что дало уникальный фосфорилируемый остаток среди дегронов семейства WNK. Эксперименты по поляризации флуоресценции и структурному моделированию показали, что его фосфорилирование аннулирует взаимодействие KLHL3 аналогично мутациям, вызывающим гипертензию. Вместе эти данные показывают, как домен Kelch KLHL3 может приспосабливаться к связыванию множественных изоформ WNK, и подчеркивают потенциальный регуляторный механизм для рекрутирования WNK3.
Заявление о конкурирующих интересах
Авторы заявили об отсутствии конкурирующих интересов.
сокращения
- BTB
- BRIC-A-BRAC, Tramtrack и широкий комплекс
- CCT
- CCCT
- COCT
- CLL3
- Cullin3-зависимое кольцо E3 Ligase
- COCT
- DMEM
- DULBECCO Модифицированная среда Eagle
- Dithiothreitol
- FBS
- FBS
- FBS
- FBS
- FBS
- Petal Bovine Serum
- FBS
- GST
- Glutathion S-Transferrate
- HCD
- HCD
- HEK293T
- Hek293T
- Embryonit Chipney 293 клеточная линия, экспрессирующая SV40
- KCC
- KCC
- K + / CL- CO-Transporters
- IPTG
- ISOPROPYL β-D-1-THIOGALACTOPIRANOSIDE
- KLHL3
- Kelch-Alke Белок 3
- LC / MS
- Жидкая хроматография-масс-спектрометрия
- N [K]CC
- Сотранспортеры ионов Na+/Cl–
- NKCC2
- Сотранспортеры Na+/K+/2Cl– 2 9155 0 OSR1
- Окислительный стресс-отзывчивый Kinase 1
- PDB
- Phaii Databank
- Phaii
- PhaiudogyPoaldostoостеронизм Тип II
- Phaii
- Кольцо
- Действительно интересный новый Gene
- SPAK
- SPS1, связанный с Proline / Alanine Kinase
- TCEP
- трис(2-карбоксиэтил)фосфин
- TEV
- вирус травления табака
- WNK
- без лизин (K) киназы
Расширение файла WNK.
 Что такое WNK.wnk а как открыть?
Что такое WNK.wnk а как открыть?Вы здесь, потому что у вас есть файл с расширением .wnk. Файлы с расширением .wnk могут запускаться только определенными приложениями. Возможно, файлы .wnk представляют собой файлы данных, а не документы или медиафайлы, что означает, что они вообще не предназначены для просмотра.
Файлы с расширением .wnk используются программой Wink. Эта программа используется для создания руководств по программному обеспечению, которые помогают пользователям легко начать использовать конкретную программу.Файлы с этим расширением обычно представляют собой файлы презентаций, созданные программным обеспечением, и обычно содержат коллекции слайдов в виде снимков экрана, сделанных программой. Эти файлы .wnk также могут содержать другие данные, такие как звук, формы, заголовки слайдов, стили, а также свойства внешнего вида. Эти обучающие файлы действительно могут помочь пользователям узнать много нового об определенном программном обеспечении. Другим приложением, которое использует файлы .wnk, является Three Degrees, приложение, позволяющее пользователям входить в приложение P2P с использованием файла .чистый паспорт. Это программное обеспечение отвечает за групповые чаты, а также другие функции мессенджера. Файлы с расширением .wnk можно открыть даже без программы Wink или программы Three Degrees, поскольку для их открытия можно использовать конвертер файлов и редактор.
Запустите файл .wnk или любой другой файл на вашем ПК, дважды щелкнув его. Если ассоциации файлов настроены правильно, приложение, предназначенное для открытия файла .wnk , откроет его.Возможно, вам потребуется загрузить или приобрести правильное приложение. Также возможно, что на вашем компьютере установлено правильное приложение, но файлы .wnk еще не связаны с ним. В этом случае, когда вы пытаетесь открыть файл .wnk, вы можете сообщить Windows, какое приложение подходит для этого файла. С этого момента при открытии файла .
Другим приложением, которое использует файлы .wnk, является Three Degrees, приложение, позволяющее пользователям входить в приложение P2P с использованием файла .чистый паспорт. Это программное обеспечение отвечает за групповые чаты, а также другие функции мессенджера. Файлы с расширением .wnk можно открыть даже без программы Wink или программы Three Degrees, поскольку для их открытия можно использовать конвертер файлов и редактор.
Запустите файл .wnk или любой другой файл на вашем ПК, дважды щелкнув его. Если ассоциации файлов настроены правильно, приложение, предназначенное для открытия файла .wnk , откроет его.Возможно, вам потребуется загрузить или приобрести правильное приложение. Также возможно, что на вашем компьютере установлено правильное приложение, но файлы .wnk еще не связаны с ним. В этом случае, когда вы пытаетесь открыть файл .wnk, вы можете сообщить Windows, какое приложение подходит для этого файла. С этого момента при открытии файла . wnk будет открываться нужное приложение. Нажмите здесь, чтобы исправить ошибки ассоциации файлов .wnk
wnk будет открываться нужное приложение. Нажмите здесь, чтобы исправить ошибки ассоциации файлов .wnkОтладочный режим подмигивания
Отладочный режим Подмигивание
Wink принадлежит к элитной коллекции программного обеспечения DebugMode и помогает пользователям легко создавать программы для презентаций или учебных пособий.В первую очередь он создан, чтобы помочь пользователям делать обучающие уроки по использованию другого программного обеспечения. Wink позволяет легко делать снимки экрана, при этом пользователь может легко добавлять поясняющие поля, заголовки и кнопки на каждую страницу. Это одна из самых простых в использовании программ для создания учебных пособий. С его помощью пользователи могут легко научить своих учеников работе с основными программами, такими как MS Word, Adobe Flash и Photoshop. Wink может легко делать снимки экрана и движения мыши, поскольку пользователи добавляют пояснения к каждому модулю.Программное обеспечение использует пользовательский интерфейс на базе Windows, который поддерживает действие перетаскивания. Все это делает создание и редактирование высококачественных учебных пособий и документации довольно простым. Wink — это бесплатное программное обеспечение, содержащее множество дополнительных функций.
Это одна из самых простых в использовании программ для создания учебных пособий. С его помощью пользователи могут легко научить своих учеников работе с основными программами, такими как MS Word, Adobe Flash и Photoshop. Wink может легко делать снимки экрана и движения мыши, поскольку пользователи добавляют пояснения к каждому модулю.Программное обеспечение использует пользовательский интерфейс на базе Windows, который поддерживает действие перетаскивания. Все это делает создание и редактирование высококачественных учебных пособий и документации довольно простым. Wink — это бесплатное программное обеспечение, содержащее множество дополнительных функций. Будьте осторожны, не переименовывайте расширение на .wnk files или любые другие файлы. Это не изменит тип файла. Только специальное программное обеспечение для преобразования может преобразовать файл из одного типа в другой.
Расширение файла — это набор из трех или четырех символов в конце имени файла; в этом случае .wnk. Расширения файлов говорят вам, какой это тип файла, и сообщают Windows, какие программы могут его открыть. Windows часто связывает программу по умолчанию с каждым расширением файла, поэтому при двойном щелчке файла программа запускается автоматически.Когда этой программы больше нет на вашем компьютере, иногда вы можете получить сообщение об ошибке при попытке открыть связанный файл.
Включите JavaScript, чтобы просматривать комментарии с помощью Disqus.Что такое файл .WNK и как его открыть — режим отладки (простые шаги)
Введение
Вы загрузили файл с расширением .wnk , но не уверены, что это такое? Скорее всего, вы знаете, что такое файл , но вы можете не знать, как его открыть. Не волнуйтесь! В этой статье объясняется все о расширении файла .wnk, как открыть и работать с ним в различных системах.
Не волнуйтесь! В этой статье объясняется все о расширении файла .wnk, как открыть и работать с ним в различных системах.
Что такое файл .WNK?
Расширение файла .WNK взято из презентации Wink . Это расширение файла было разработано DebugMode . Формат этого файла представляет собой двоичный файл , и это файл, связанный с данными.
Что нужно знать
В настоящее время существует 1 программа, которая может открывать файлы .wnk на 2 платформах.
Как открыть файл .WNK?
Ниже мы написали подробное руководство о том, как открыть файл .WNK в различных операционных системах.
Как открыть файл .WNK в Windows (7, 8, 10)
Ниже мы написали простое для понимания руководство по , как открыть файл .wnk на Windows за 4 простых шага.
1. Первым шагом является загрузка программного обеспечения DebugMode Wink.Вы будете использовать это программное обеспечение, чтобы открыть файл . Другое программное обеспечение, которое вы можете использовать, — это DebugMode Wink.
Шаг 2. Поиск файла .WNK
2. Второй шаг — найти загруженный файл . Файл обычно можно найти в папке /download/ , если вы не помните, куда вы его загрузили.
Шаг 3. Выберите «Открыть с помощью»
3. На третьем шаге, найдя файл , щелкните его правой кнопкой мыши и выберите «Открыть с помощью» .
Шаг 4: Выберите программное обеспечение и нажмите «ОК»
4. После выбора опции «Открыть с помощью» у вас будет возможность выбрать загруженное вами программное обеспечение DebugMode Wink . Нажмите на программное обеспечение и нажмите «ОК» . Поздравляем! Вы открыли свой файл на Windows .
Нажмите на программное обеспечение и нажмите «ОК» . Поздравляем! Вы открыли свой файл на Windows .
Как открыть файл .WNK в Linux
Чтобы открыть файл .wnk на Linux , выполните следующие 4 простых шага.
1. Первым шагом является загрузка программного обеспечения DebugMode Wink. Вы будете использовать это программное обеспечение, чтобы открыть файл . Другое программное обеспечение, которое вы можете использовать, — это DebugMode Wink.
Шаг 2. Найдите файл .WNK
2. На втором этапе вам нужно найти загруженный файл .Если вы не помните, куда вы загрузили файл , вам следует проверить папку /download/ , так как обычно он автоматически сохраняется там.
Шаг 3. Наведите указатель мыши на кнопку «Открыть с помощью»
3. На третьем шаге щелкните правой кнопкой мыши файл и выберите «Открыть с помощью» .
На третьем шаге щелкните правой кнопкой мыши файл и выберите «Открыть с помощью» .
Шаг 4: Откройте файл, выбрав программное обеспечение
4. На четвертом этапе после нажатия «Открыть с помощью» вы должны увидеть загруженное вами программное обеспечение DebugMode Wink .Выберите свое программное обеспечение и нажмите «ОК» . Вот и все, теперь у вас открыт ваш файл на Linux .
Все еще не можете открыть файл .WNK?
Если вы по-прежнему не можете открыть файл .WNK , это может быть связано с тем, что вы пытаетесь открыть неправильный файл. Расширение файла .WNK похоже на другие расширения файлов , такие как: .wnry — файл, зашифрованный вирусом WannaCry, .woa — Apple, .woff — рабочая группа WebFonts, .